|
|
BIOL
4120
Principles of Ecology
Phil Ganter
320
Harned Hall
963-5782
|
The
dead arm of the cactus above is home to populations of yeast,
bacteria and insects but is a habitat that persists only for a
short time. How do species that live in temporary habitats
persist? |
Lecture 12 Metapopulations
Email me
Back to:
Overview - Link
to Course
Objectives
Metapopulations
A region may contain more than one
population of any given species. A Metapopulation
is the larger unit made up of a group of Subpopulations
(local populations of one species).
- Several organizational schemes
for metapopulations exist:
- Patchy Populations
- A group of subpopulations interconnected
through migration
- Some subpopulations might go extinct,
but the metapopulation persists and can recolonize empty patches
- Metapopulation is stable through
constant extinction-recolonization events
- an equilibrium is reached
which balances local extinction and recolonization
- for this kind of metapopulation,
the regulation of the metapopulation is not directly tied to the fate
of any one subpopulation
- Core/Satellite
or Mainland/Island or Source/Sink
metapopulations (terms are equivalents)
- A large population with smaller,
satellite populations founded from the big one
- Notice that the large population
is often treated here as a source (and is relatively unaffected
by the satellites) and the satellites are affected by the core,
but not by one another
- for this kind of metapopulation,
the regulation of the core population drives the long-term fate of
the satellite populations
- Rescue
Effect - This concept
is usually applied in Island Biogeography, a subset of metapopulation
theory.
- When an island is close to its source
of species, both the rate of colonization and the rate of extinction
are affected
- Colonization is affected by the
proximity of the mainland
- The rescue effect is a reduction
in the extinction rate because a population on an island is
replenished or re-colonized from the source (the mainland)
- The rescue effect comes from
the arrival of new members into a population already established
on the island
- These new arrivals increase
the population size and decrease the probability that
the population will go extinct.
- Isolated, Nonequilibrium
Populations
- Like patchy populations, but there
is no migration between patches so that once extinct, a subpopulation
is lost
- if the populations
do not exchange migrants, each is a closed system and the population
growth models we have discussed describe their dynamics
- the changes in the
species in the region would be the sum of changes in the independently
changing populations in the region
- Metapopulation is unstable, as subpopulations
are lost but no new ones are established
- for this kind of metapopulation,
the possibility of local extinction implies the possibility of metapopulation
extinction
- Habitat
Fragmentation by human activity can create metapopulations
with isolated subpopulations from previously large local populations
- fragments left by
human activities may be too small to sustain breeding populations
- fragments may be
too far apart to allow sufficient migration
- Checkerspot butterfly
(Euphydryas editha) is an example of what can happen
to species whose habitat has been affected by human activity
- large source population and nine
satellites (all on Jasper Ridge)
- present on Jasper Ridge since at
least 1934
- In 1996, core population collapsed
and eventually the species disappeared from Jasper Ridge
- satellite populations did
not persist long after core was lost
NOTE: The book covers a famous example of insect metapopulations, the
populations of the bay checkerspot butterfly, Euphydryas editha, near
San Francisco, CA. The populations occur on ridges composed of serpentine
soil. This does not mean soil with lots of snakes. Serpentine soil
is named for its parent rock, a group of minerals (collectively called serpentine
minerals) that are hydrous iron magnesium phyllosilicates. There are over
20 minerals in this group and the rocks made of mixtures of these minerals are
called serpentinite. They also contain other metals including chromium
and cobalt. The rocks are often greenish and, when polished, must have
reminded someone of a snakeskin (hence the name may refer to snakes).
They are igneous rocks (actually, ultramafic) that are from the Earth's mantle
and tend to occur at the surface as intrusions into the continental rock, which
is the reason for the patchy occurence in the San Francisco area as ridges of
serpentinite rock. For our purposes, serpentine soils contain high percentages
of serpentinite and the soil moisture will have high concentrations of the metals
characteristic of the rock. These metals are toxic at such high concentrations
and many plants can not grow on serpentine soils. Those that do
occur there often occur only on serpentine soils, which are not common and so
the plants found there are often rare and appear on lists of threatened or endangered
species.
Extinction
and Colonization
Patchy populations are of interest to ecologists
because the metapopulation may persist even thought the local populations are
guaranteed to go extinct. The persistence of the metapopulation is truly
an emergent property!
To see how this happens and to explore
the dynamics of this situation, we will use a mathematical model. The
model will describe the outcome of the operation of two processes:
- Extinction:
the loss of a local population for any reason (disease, visit by a predator,
demographic stochasticity, etc.). This is sometimes called Local
Extinction because it refers to local populations.
- Colonization:
The founding of new local populations from existing populations
A
Model of Patchy Metapopulations
Before building the model, we
need to state the assumptions built into the model
- This model applies to patchy
metapopulations, which implies that the habitat consists of discrete patches
of suitable habitat surrounded by unsuitable habitat and that there is some
degree of dispersal between patches. From this second assertion, we
can infer that suitable patches which are not occupied by a local population
are subject to colonization from occupied patches.
- Each local population has a
non-zero chance that it will go extinct in a specified time interval
- The changes in local populations
are not synchronized or correlated.
Now we can build the model and
analyze its output. Which output is of interest? Metapopulation
size is one possibility but predicting size might be complicated as we would
need to predict subpopulation sizes. We can simplify local population
size to one of two states: occupied or not occupied. This gets
rid of lots of complications but we must assume that once occupied a patch
puts out migrants at some constant rate until it becomes extinct. With
this simplification, we can focus on a simplified metapopulation size: the
proportion of patches in a region that are occupied. As it increases,
metapopulation size increases. Suppose we call P
the proportion of patches that are occupied by a local population of the species
of interest and e is the probability
that a patch will go extinct within a specified time. The rate of extinctions
we expect during the time interval is :
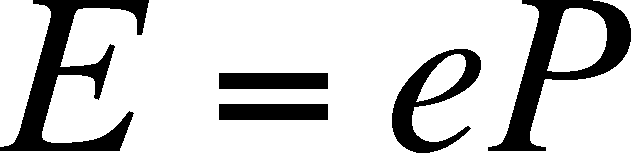
The colonization rate depends
on a probability of a migrant finding a patch, m, and the P, the proportion
of patches that are occupied. P affects the colonization rate in a direct
and an indirect way. The direct effect is that, as P increases, the
number of dispersing individuals increases and indirectly because, as P increases,
the number of unoccupied patches decreases and the chance of finding such
a patch decreases. Thus, the colonization rate is:
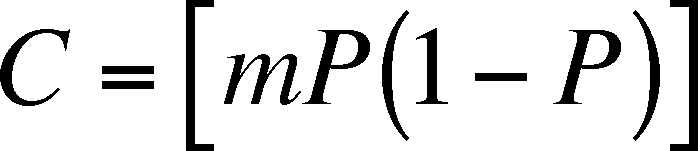
The rate of change in the proportion
of patches occupied (DP/Dt)
is the difference between the colonization rate and the extinction rate (=
C - E)
or, with a little substitution:
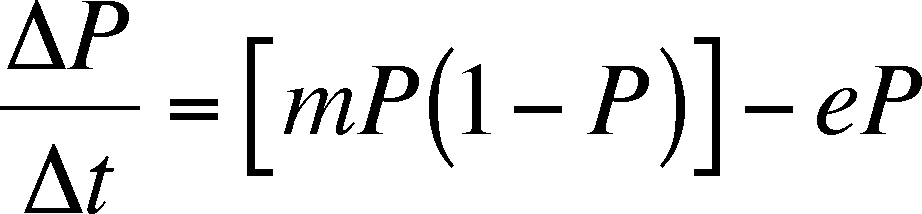
For any given probability of extinction
and probability of colonization (m)
and extinction (e), we can plot
the extinction (E) and colonization
(C) rates versus the proportion
of occupied patches (P).
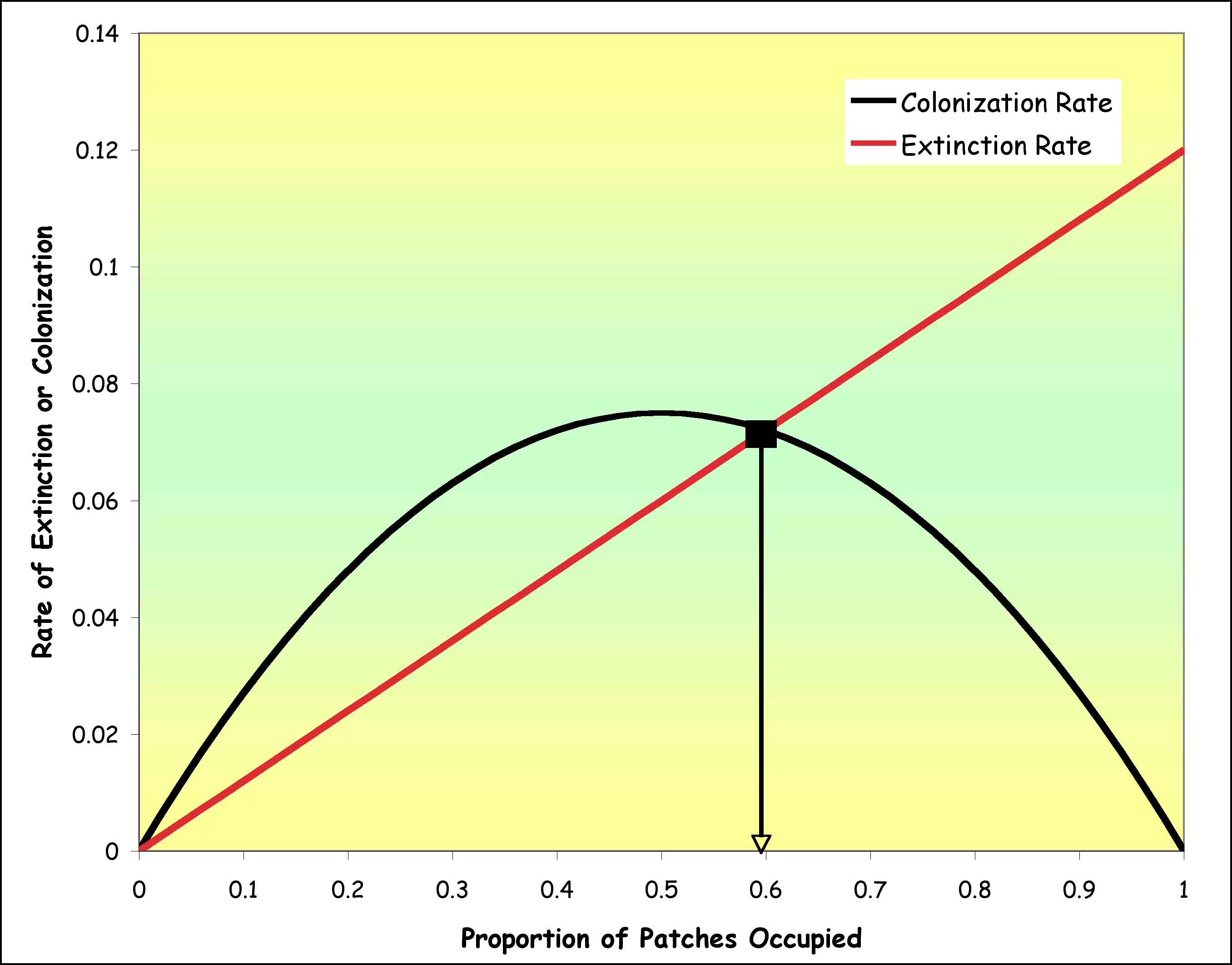
The extinction rate changes linearly
(the more patches that are occupied, the more that go extinct) but the colonization
rate is humped, with the highest values found in the middle, where neither
the patches providing dispersers nor unoccupied patches are rare.
Where the lines cross, the extinction
rate equals the colonization rate and there is no net change in the proportion
of patches occupied (for each loss a new population is begun). We can
see that this does not have to happen (imagine a situation where the colonization
hump is very low and the extinction line is very steep. What determines
where the two lines cross? That is the point where the rate of change
in P (DP/Dt)
is zero. To find this, we set DP/Dt
= 0 and we solve the equation to find that the rate of change in P
is zero when:
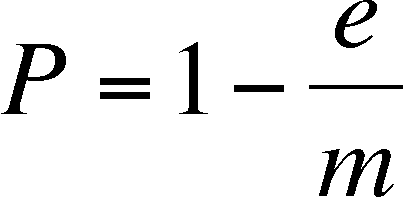
So, we have a prediction for how
the rates of extinction and colonization can produce a stable metapopulation
with a predictable proportion of patches occupied. Note that the equation
above will produce nonsense if we allow e to exceed m (we would get a negative
proportion of patches occupied, whatever than means). Thus, the model
needs to specify that m >
e if the metapopulation is to
persist. We will use this relationship below to understand how changes
in m and e
affect the proportion of patches occupied (P)
Patch
Size and Isolation
Patches can change on many ways.
Some are high quality while others may be of lower quality. Some
may be large and some may be isolated. We can explore how these changes
might affect the metapopulation with the model. We will focus on two factors:
patch size and patch isolation.
How would a change in patch size
affect metapopulation dynamics?
- Large patches should lead to
large populations. Large local populations should have a smaller chance
of going extinct through due to demographic stochasticity. This might
even be true for more specific reasons for extinction. Large populations
would be more likely to have individuals resistant to a disease. Large
populations may exceed the ability of a predator to drive the local population
to extinction.
- There is no clear relationship
between population size and colonization rate.
How would Isolation (distance between
patches) affect metapopulation dynamics?
- Since extinction rate is a local
phenomenon, isolation will not affect it.
- Greater distance between populations
will affect the colonization rate by making it more difficult to find unoccupied
patches.
These two forces will then change
the proportion of patches occupied. A diagram will make the differences
easier to understand. We can compare small to large patches by comparing
the extinction rate line produced with a small e (= large populations) to a
line produced by a larger e (= small populations). On the same graph,
we can compare isolated patches (= small m) with a metapopulation with close
patches (= large m) and see how these things interact.
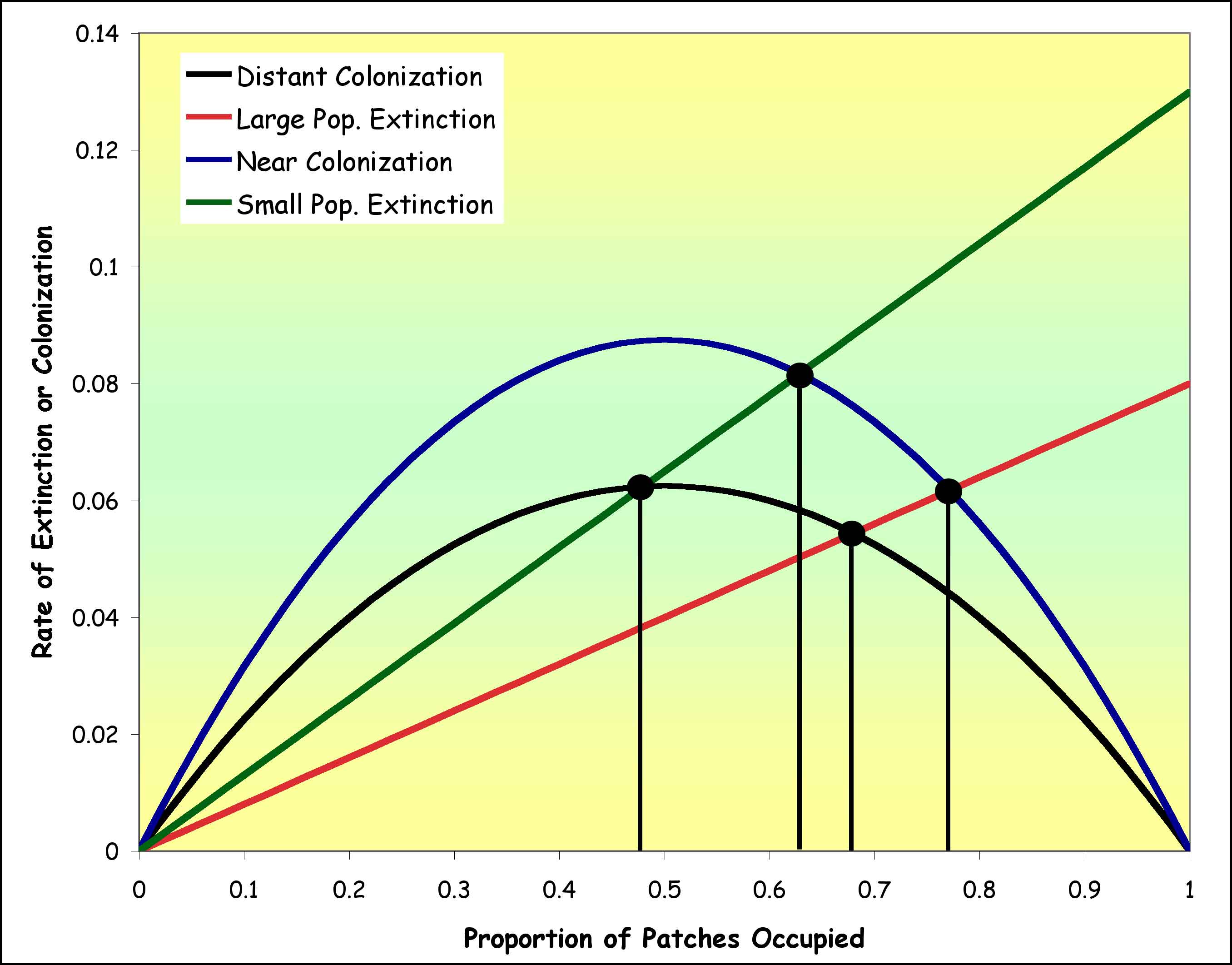
You can see that a metapopulation
with large, close patches will have a higher rate of patch occupancy than will
a metapopulation with small, isolated patches.
- Think of what such a conclusion
might mean to a resource manager. As development reduces patch size
and increases patch isolation, the possibility of metapopulation extinction
increases.
Remember that the comparisons above
will work for any factors that increase or decrease m or e. Patch size
and isolation are only examples of factors that affect m and e.
Heterogeneity
variation in
habitat quality over space (Spatial Heterogeneity)
or time (Temporal Heterogeneity)
can increase the probability of extinction
Weather is a factor which has
great heterogeneity and it only takes one severe storm, severe drought or
severe cold spell to doom local populations
In January, 2007, northern
Florida was host to severe storms that generated tornados. The storms
killed 17 of the 18 whooping cranes (Grus americana) that had
just established a winter population there. These cranes are endangered
and there is an intensive program to rescue them from extinction.
The species is migratory and overwintered in the southern US from Texas
to Florida at one time. Hunting and habitat alteration caused local
extinctions. They were almost extinct from nature (only 21 wild
birds in 1941) but a breeding program in zoos preserved the species.
There are now over 300 in the wild and about 150 in zoos. The young
in the release program were kept innocent of human contact (keepers donned
crane costumes when in the crane compound). A successful population
that breeds in Canada and overwinters in Texas has been established.
However, the conservation ecologists in charge of the project realized
that a single event in Texas or Canada could be disastrous and wanted
to establish a second population. A second breeding population is
now being attempted in Wisconsin with overwintering in Florida.
In 2001, a breeding population in Wisconsin were lead to overwintering
sites in Florida by leading them with ultralight aircraft disguised as
cranes. In Fall of 2006, they lead 18 yearling birds to Florida
to establish a second overwintering site but the prediction of disaster
turned out to be too real. Metapopulation ecology has moved out
of academia and into the real world.
- Spatial heterogeneity
is the degree to patches vary in both quality and the arrangement of
those patches (distance between)
- Each subpopulation's maximal
size depends on it's patch quality
- Spatial heterogeneity important
as a means of unlinking the fates of individual patches
- this effect is called
Spreading the Risk
- can lead to stable metapopulations
even though local subpopulations have a high probability of going
extinct
- Importance of spatial heterogeneity
to stabilizing metapopulations has been demonstrated for laboratory systems
- Huffaker demonstrated
the importance of spatial heterogeneity in simplified predator-prey
system
- studied a predator-prey system consisting
of a herbivorous mite (feeding on oranges) and a predacious mite
that ate the herbivorous mite
- on one orange, the predator
always eliminated the prey and then died off itself
- when an array of oranges
was used, mite populations on each orange eventually suffered
the fate of the populations in the single orange system
- the extinction process
took longer but still local extinction lead to metapopulation
extinction
- Huffaker increased spatial
heterogeneity
- made the system larger
(more oranges)
- made some patches
suitable (oranges) and some unsuitable (rubber balls)
- made migration more
difficult with Vaseline barriers between oranges (that
the predator could cross, although with difficulty)
- herbivores used sticks
in oranges as launch sites for "ballooning"
to next orange
- increased heterogeneity resulted
in persistence of both predator and prey
- also resulted in
predator-prey cycles!
- microbial predator-prey
systems also demonstrated that increased spatial heterogeneity stabilized
metapopulation size
Species
Characteristics
It is to be expected that metapopulation
dynamics are affected by the characteristics of the species under consideration
and that the reverse is also true. Species characteristics are affected
by metapopulation dynamics.
the r-selected species vs. K-selected
species distinction (or the ruderal - non-ruderal distinction drawn by Grime)
reflects the importance that dispersal can have on general life history characteristics.
Ruderals and r-selected species both have life history characteristics linked
to inhabiting ephemeral (persisting for only a short time) habitats, like forest
openings caused by a fallen canopy tree or in habitats like those my yeast and
flies inhabit (see picture at the top of the page).
Some examples of the interactions
among species characteristics and metapopulation dynamics:
- High fecundity and high dispersal
rates are often correlated.
- Dispersal is dangerous and
many young are needed if all disperse into a hostile environment.
- Population size is correlated
with body size, so that local populations are smaller for a species with a
large body size compared with a similar species that has a smaller body size.
- Population size is inversely
correlated with the probability of extinction and will affect metapopulation
dynamics for this reason.
Overview
of Populations
We have examined populations from
more than one perspective and it is good to integrate these varying views.
This integration can be done by viewing populations hierarchically.
- Local populations consist of
interacting individual organisms (whether modular or not) are the lowest level
of the population hierarchy.
- Individuals experience similar
conditions within the local population and so it is reasonable to summarize
their response to these conditions in a Life Table, from
which we can calculate such important population parameters as birth and
death rates and population growth rate.
- In an ideal situation, all
individuals would occupy a single patch of suitable habitat. While
this is often so, real populations may not always conform but the
assumption that the members of the population experience similar conditions
must be met.
- The next level of the hierarchy
is the metapopulation, where dispersal links local populations.
This linkage gives rise to dynamics at the metapopulation level that cannot
be seen at the local level (another of those emergent properties).
- Metapopulation dynamics are
affected to the degree to which changes in local populations are correlated
from population to population. As the correlation weakens, metapopulation
dynamics become less and less predictable from the changes occurring in
any single local population.
- Because metapopulations are
composed of populations that exchange individuals, however rarely, the
gene flow will, under most conditions, be sufficient to promote genetic
similarity among the local populations that comprise the metapopulation.
- The next level is the Subspecies.
This is one or more metapopulations that are separated from other metapopulations
of the same species by sufficient distance and/or experience sufficiently
distinct conditions that gene flow is not sufficient to maintain genetic similarity.
- There are two ways for this
isolation to occur.
- If Geographic
Barriers separate metapopulations, the boundary between
subspecies can be abrupt.
- If there are no effective
geographic barriers but the distance between the farthest metapopulations
is great, gene flow may be slow enough to allow divergence between
subspecies, even if there is no gap between metapopulations.
This phenomenon is called Isolation
by Distance.
- The book has an
example of such in the discussion of Ensatina escholtzii,
a salamander from the west coast, found in chapter 2.
- Subspecies
may diverge through the spread of different adaptations
linked to local conditions or may diverge through genetic drift
if population size is small enough and/or there is sufficient time for
divergence.
- Not all species are organized
into subspecies
and, so, this level of population organization does not always exist.
- The Species level, the final
level of population organization, consists of all local populations that can,
with some reason, be considered as members of the same species.
- The geographic range here
is often large and those who study entire species often consider their
efforts properly placed in the subdiscipline of Biogeography.
Asexual organisms, although they
can not recombine and the concept of a gene pool does not apply, can be understood
using the hierarchy above.
- Local populations, because of the requirement
that individuals within the populations experience similar conditions, are
kept genetically similar not by the spread of successful genes but by the
replacement of less successful genotypes by more successful genotypes
- Metapopulations experience genotype flow, not
gene flow, and are kept similar through the same means.
- Subspecies may arise under the same conditions
as for sexual species
- The problematic level may be the species level
of population organization when there is no obvious criterion for deciding
when differences among subspecies have become large enough to consider them
to be new species.
- If you have a solution for this conundrum,
contact me. You will be famous.
Terms
Metapopulation, Subpopulation, Patchy Population,
Core/Satellite, Mainland/Island, Source/Sink, Rescue Effect, Isolated, Nonequilibrium
Populations, Habitat Fragmentation, Extinction, Local Extinction, Colonization,
m, e, E, C, P, DP/Dt,
Spatial HeterogeneityTemporal Heterogeneity, , Subspecies, Geographic Barriers,
Isolation by Distance, Biogeography
Last updated February 10, 2007
