|
BIOL 4120 Principles of Ecology Phil Ganter 320 Harned Hall 963-5782 |
Lecture 14 Predation/Herbivory
Back to:
| |
BIOL 4120 Principles of Ecology Phil Ganter 320 Harned Hall 963-5782 |
Lecture 14 Predation/Herbivory
Back to:
Overview - Link to Course Objectives
An interaction between two species in which one is harmed and the other helped
Several flavors of interaction fall under the above rubric:
- Herbivory -- plant eaters, algae not usually considered but included
- Carnivory -- meat eaters (other carnivores or herbivores)
- Cannibalism - eating one's own species - a specialized form of predation
- Parasitoids -- usually insects that lay their eggs on other insects as hosts. The larvae complete development on the host, usually killing the host as a result
- Parasitism -- feeding on another organism's parts without killing the organism (a note: parasitism is very widespread and includes all kingdoms as both parasite and host)
The chart below can be helpful in separating the types (adapted from text)
| Lethality to host |
||
| Contact between organisms |
Low |
High |
| Close and Long-term | Parasites |
Parasitoids |
| Brief |
Herbivores |
Predators |
Both predation and herbivory are +.- species interactions in which one species benefits from the contact and one is harmed.
- Predation is a form of carnivory in which food items (prey) are searched for (or waited for), caught and killed.
- Predators tend to be much larger than prey to minimize the risk to the predator (predators do not give the prey a "fighting chance" and there is very little "struggle" in the struggle for life)
- Parasitoids are insect predators differ from the standard model of predation.
- The adult searches for and captures the prey.
- Eggs are then laid on the prey.
- The larvae hatch and eat the prey, pupate, and emerge as adults to search for prey for their offspring.
- Herbivory is a bit the same as and a bit different from predation.
- It involves plants.
- Herbivores may be much larger (cattle eating grass) or much smaller (caterpillars eating leaves on a tree) than their food items.
- Herbivores must also search for food items.
- The food items are usually not killed but are damaged by the herbivore.
- If the herbivore eats the seeds, then the plants are killed and some large herbivores do kill the plants they attack (elephants often knock down trees to get to the leaves, cattle and sheep sometimes pull out the plant by its roots).
- Predator-prey interactions are usually of short duration.
- Some herbivores spend the rest of their lives on the plant once it is found (sometimes multiple generations).
A better way to look at this might be to divide herbivores into those that are like predators and those that are like parasites
- Predator-like herbivores are larger than their food items, the encounter between herbivore and plant is of short duration compared to length of herbivore's life, and the encounter may kill the plant.
- Many vertebrate herbivores might be in this group,
- Parasite-like herbivores are smaller than their food items, the herbivore-plant encounter lasts for a long time, and the plant is usually not killed.
- Many insect herbivores are in this category.
We will use an approach that builds on the logistic that we have worked with previously
First, we assume that the prey population will grow exponentially except that the predator is there to eat some of the prey
If we set N as the number of prey, C as the number of predators, and use other, familiar terms, then the rate of change of prey is:
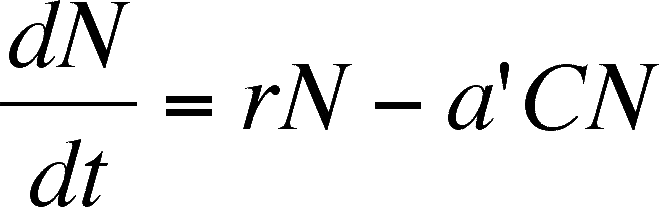
Read this equation as: The rate of change in the number of prey present (dN/dt) is a function of the birth of new prey (rN) minus the death of others due to predation (a'CN). The death rate is assumed to depend on the number of prey, the number of predators (C) and the fudge factor (a').
The rate of change in the predator population is:
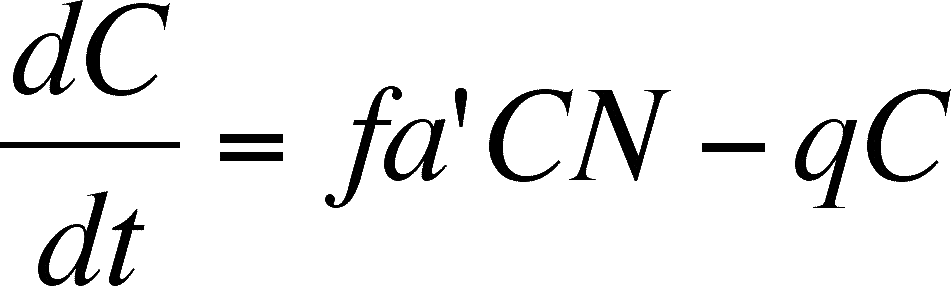
Read this as: The change in the number of predators present is a function of the birth of new predators minus the death of others due to some constant mortality rate (old age? mishap? parasitism?). The birth rate is assumed to depend on the number of prey (the resource the predator uses to make new offspring), the number of predators (more predators, more predator offspring), a' (the ability to catch prey), and a second fudge factor called f, which is the efficiency with which predators turn their food (prey) into offspring.
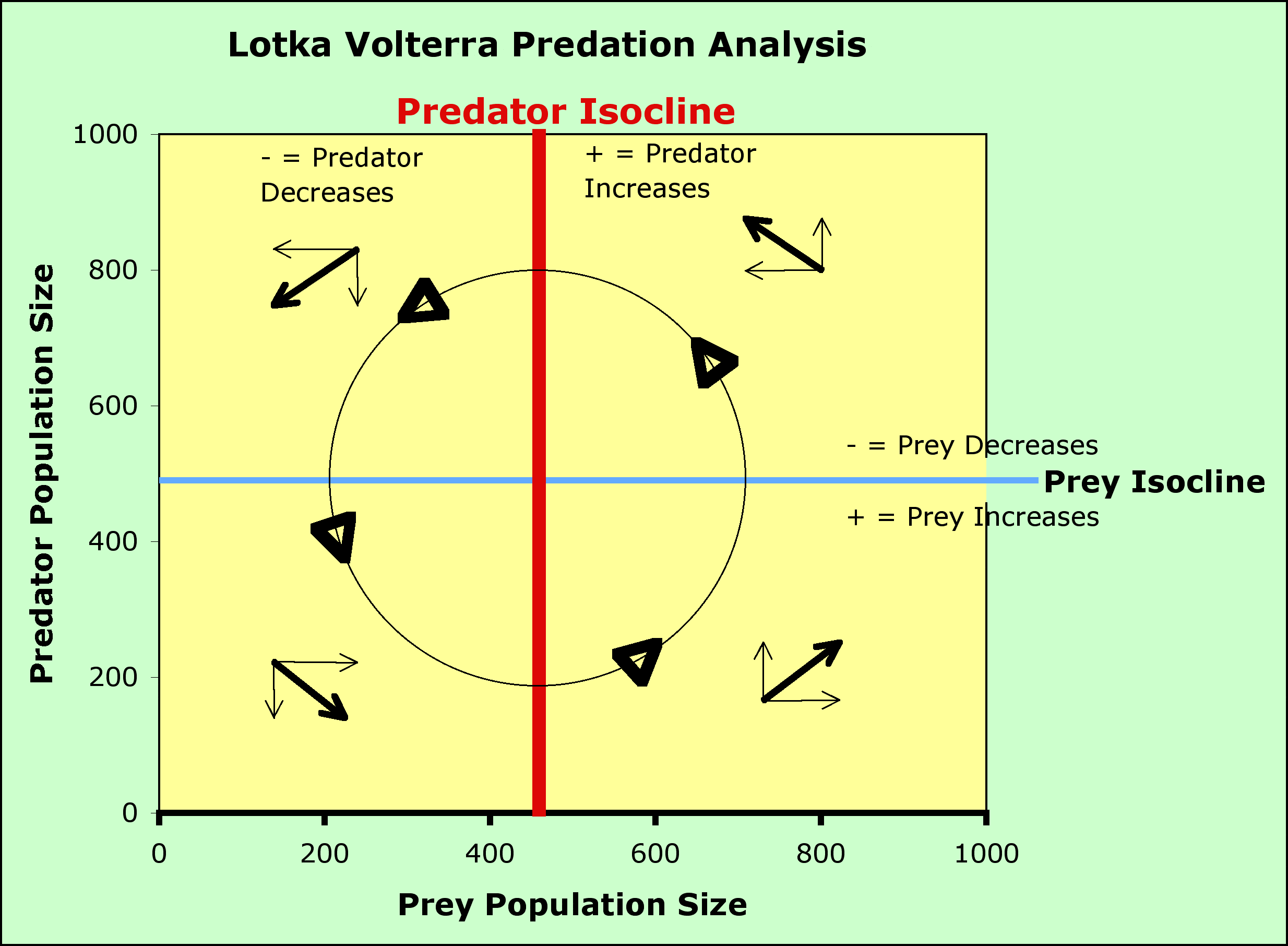
Taking the same tack as before, we solve these equations for conditions where no growth is taking place (hence some sort of equilibrium) by substituting 0 for dN/dt and for dC/dt:
Prey zero isocline:
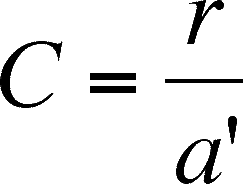
So, prey numbers do not grow when the number of predators is equal to the ratio of the prey's intrinsic rate of increase (a property of the prey) and a', the searching efficiency of the prey-predator combination (the graph of this is a straight line parallel to the prey axis - usually the x axis). When the number of predators is below this line, the prey increase, when the predator number is above the line, the prey decrease.
Predator zero isocline
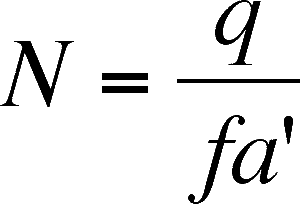
So, predator numbers do not grow when the number of prey is equal to the ratio of the predator's intrinsic death rate (a property of the predator) and the product of f and a', (this is also a straight line but is parallel to the predator axis, not the prey axis - usually the y axis). When the number of prey is to the left of this line, the predators decrease (not enough prey per predator). When the number of prey is to the right of this line, there are sufficient prey per predator and the number of predators increases.
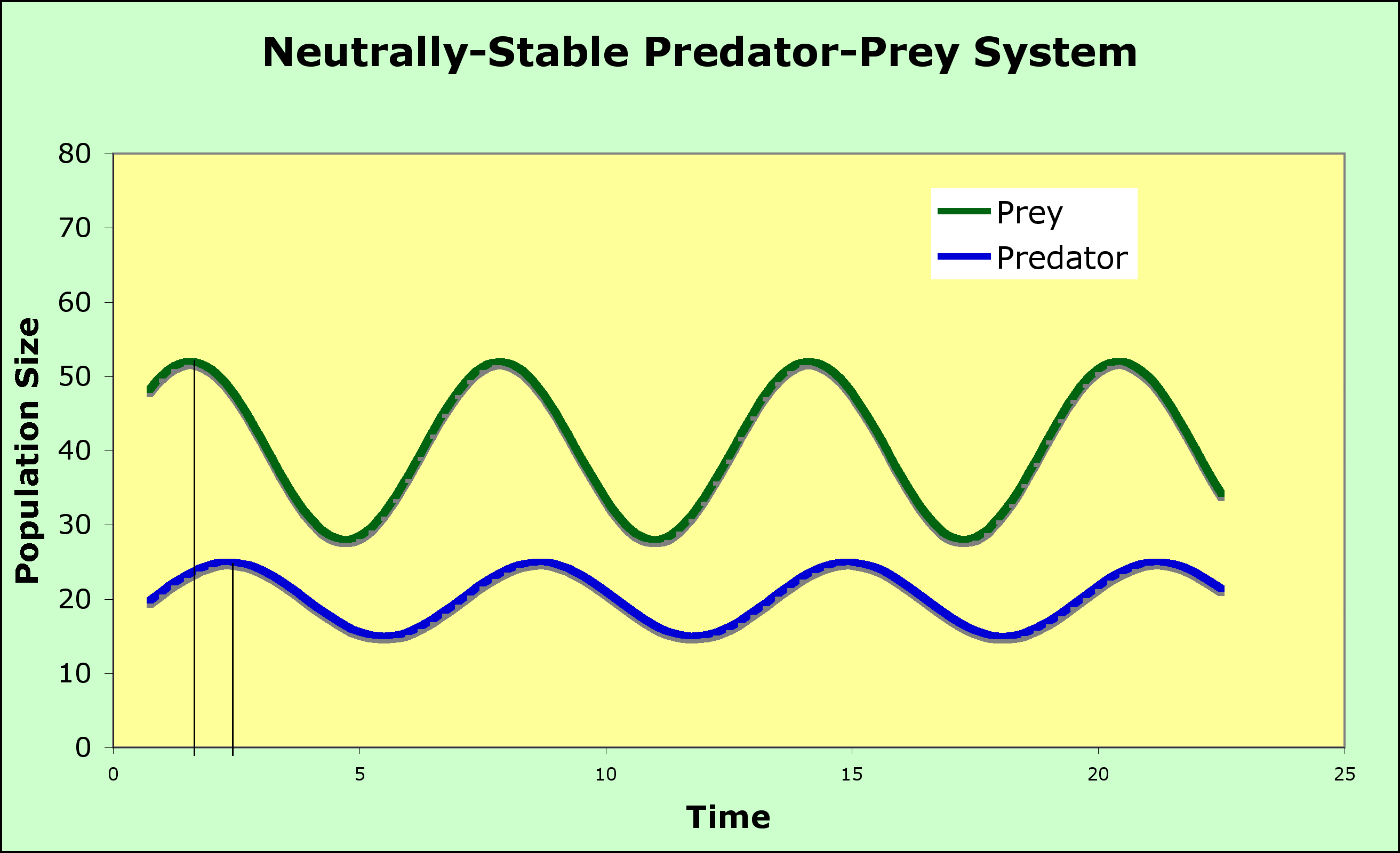
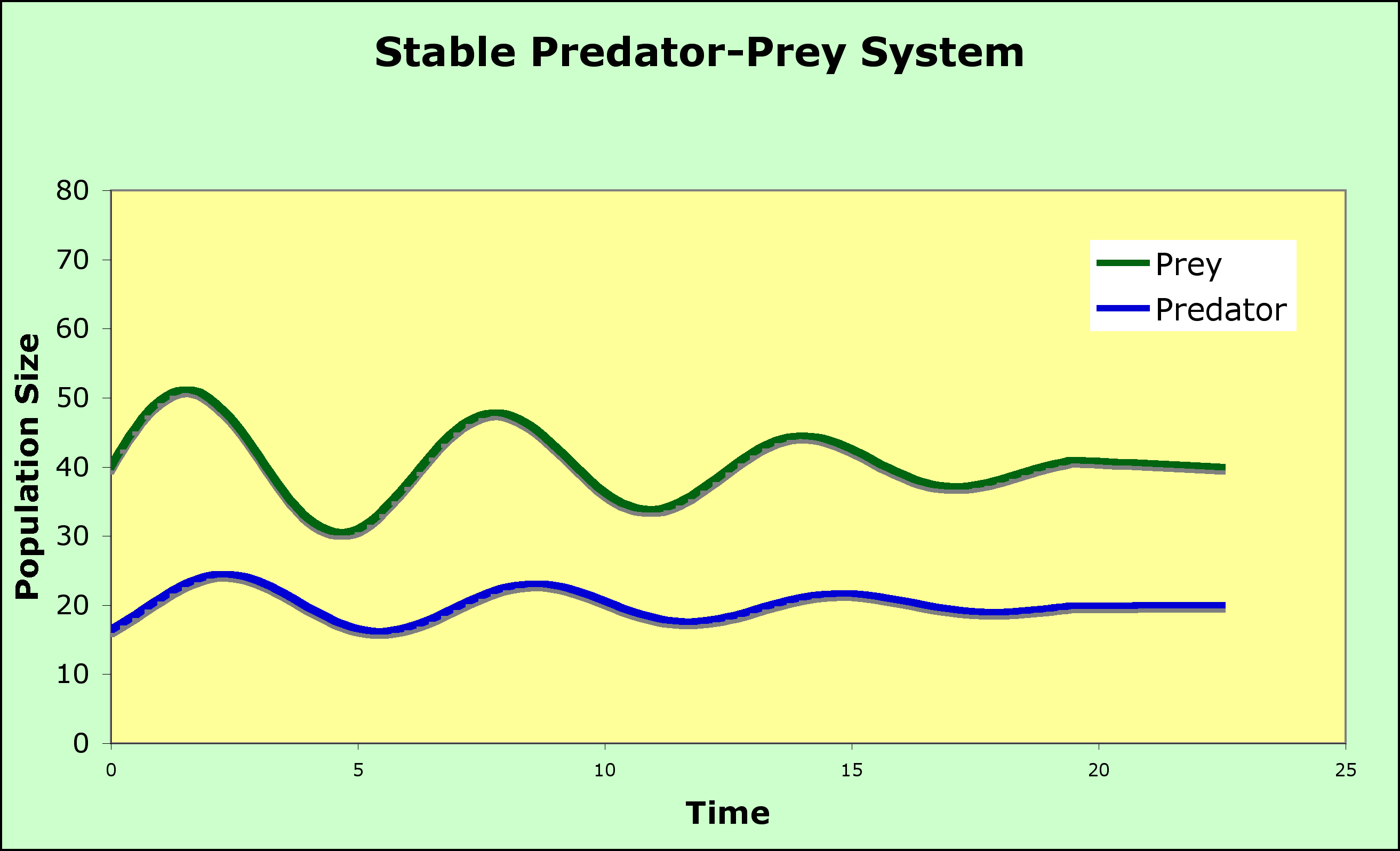
This situation leads to an Equilibrium (where the isoclines cross).
- The blue and red lines are the zero growth isoclines
- Look at the + and - regions for predator and prey on the graph - be sure you understand why above the blue line the prey population decreases and why the predator population decreases to the left of the red line.
- Look at the vector addition arrows - be sure you can explain the direction of each set of arrows
- Look at the circle and the arrowheads on the circle - be sure you can explain why the arrow heads indicate counterclockwise motion.
- Notice that there are no other equilibrium points on the axes except for 0,0 (the origin) where neither are present
- With no prey present, the predators decline to 0
- With no predators present, the prey population takes off to infinity (exponential growth, as we mentioned at the beginning of the modeling effort in this chapter)
- The second equilibrium point is where the isoclines intersect
- These equations are neither stable nor unstable (this condition is called neutrally stable), as the system will oscillate in what is called a predator-prey cycle - see graph below
- prey increase until there is enough for predators to start to increase
- predators increase until they eat enough prey to cause a decline in the prey population
- prey start to decline in number until predators can't find enough to eat and the predator population declines
- prey begin to increase, and we are back at step 1
- Some evidence for these under special circumstances
- Lynx and hare
The predictions of the model can be modified by changing the shape of the isoclines to include:
A Graphical model of Predation
Now that you are used to a graphical analysis of species interactions, a completely graphical analysis of predation first proposed by Michael Rosenzweig and Robert MacArthur can be used to stabilize the model above
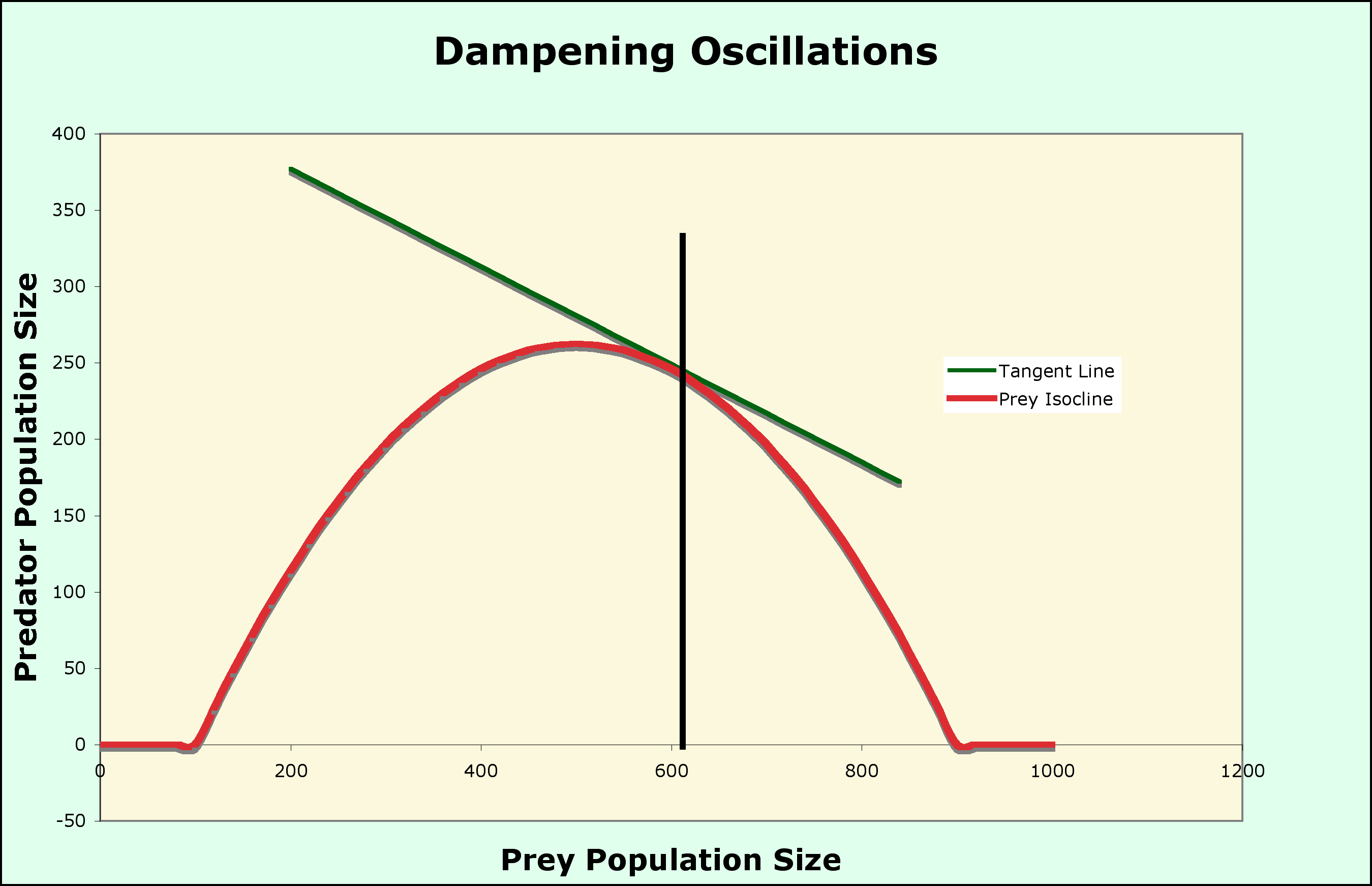
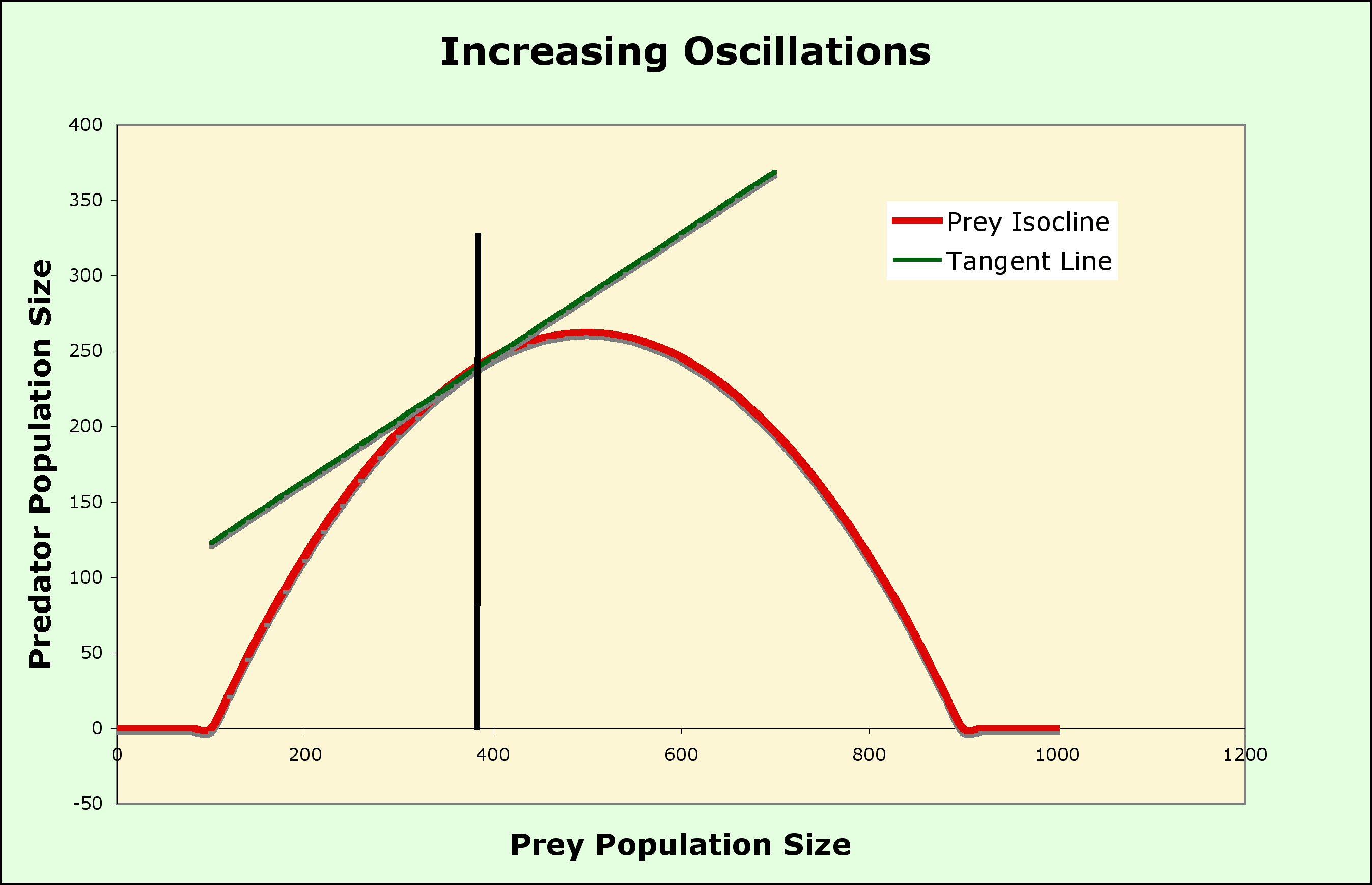
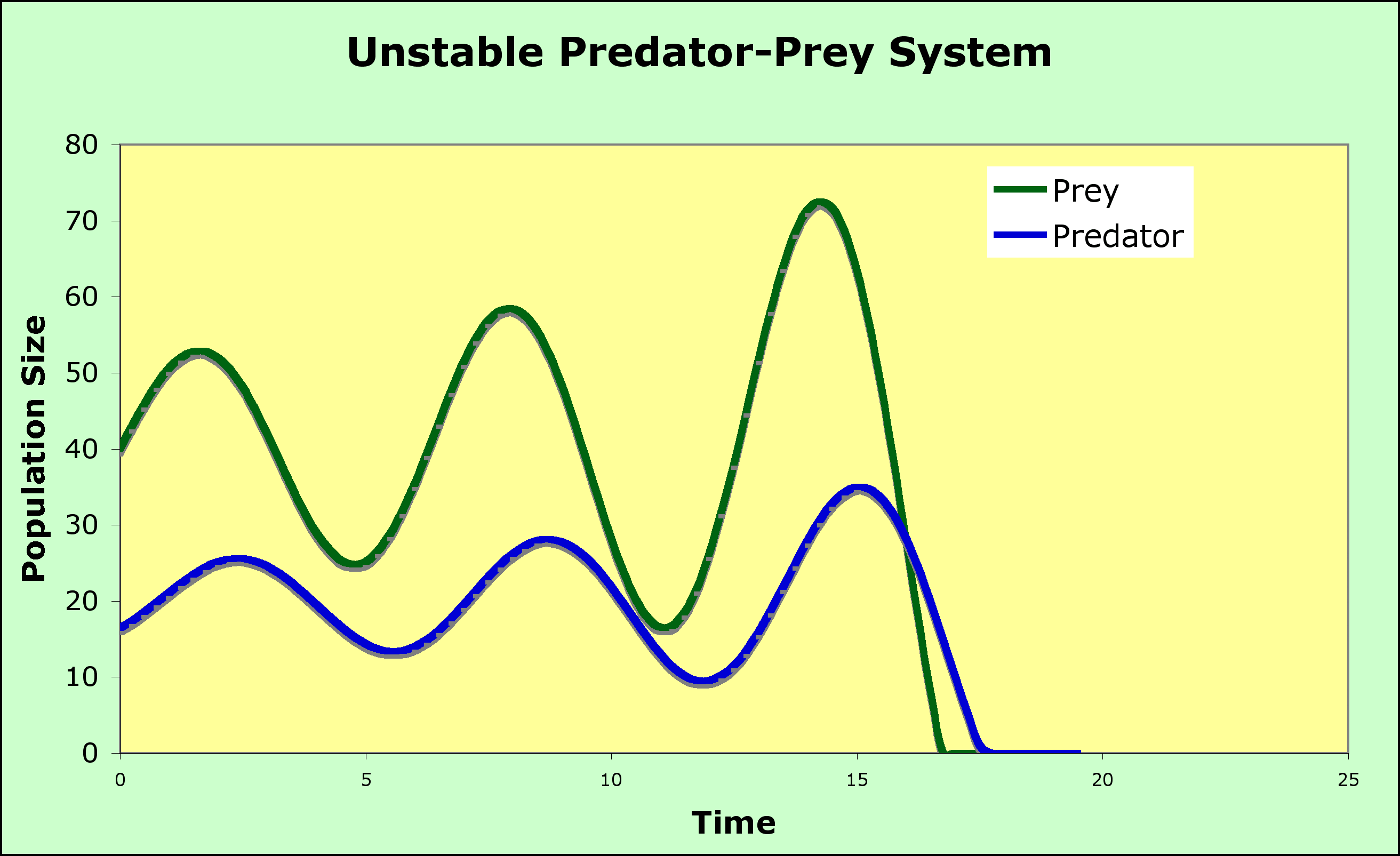
Two variations
The paradox is that, by enriching the environment, you destabilize the system! Not all good acts have the intended outcome.
Predators with more than one available prey type must decide which prey to pursue. Optimal Foraging theory has been developed to understand the factors that govern this decision for any organism with more than one choice of food.
- This theory applies to any situation in which there is a choice between food of different quality (different types, large versus small items of food of the same type, high versus low quality food items of the same type).
Optimal Foraging is based on some assumptions.
- The forager must make its choice based on maximizing its intake of some aspect of its food (calories, an essential nutrient, etc.).
- There must be a cost to each choice. This cost can be the difficulty of "handling" the food, ingestion of a toxin found in the food, the time it takes to search for the food, the chance that searching will expose the forager to a predator, or some other cost.
Optimal foraging is another example of a Trade-Off based on costs and benefits.
- Optimal forager will choose prey to maximize the benefit/cost ratio.
- This often means that the optimal strategy is to avoid a choice prey item if the costs are too great.
How should a predator find its prey?
- Some predators are Sit-and-Wait Predators. Here the search strategy involves the choice of a place in which to wait.
- Active, hunting predators search in a heterogeneous environment composed of patches with prey, patches without prey, and some distance between patches.
The Marginal Value Theorem (from economics) predicts the length of time a predator should spend in a patch based on the distance between patches, the rate of prey discovery within the current patch (more elaborate versions of the model include variation among quality of patches).
- The best strategy is the one that will maximize the predator's total intake of some aspect of its food (total prey caught, total calories, total of an essential nutrient, etc.) over the total search time available to a predator.
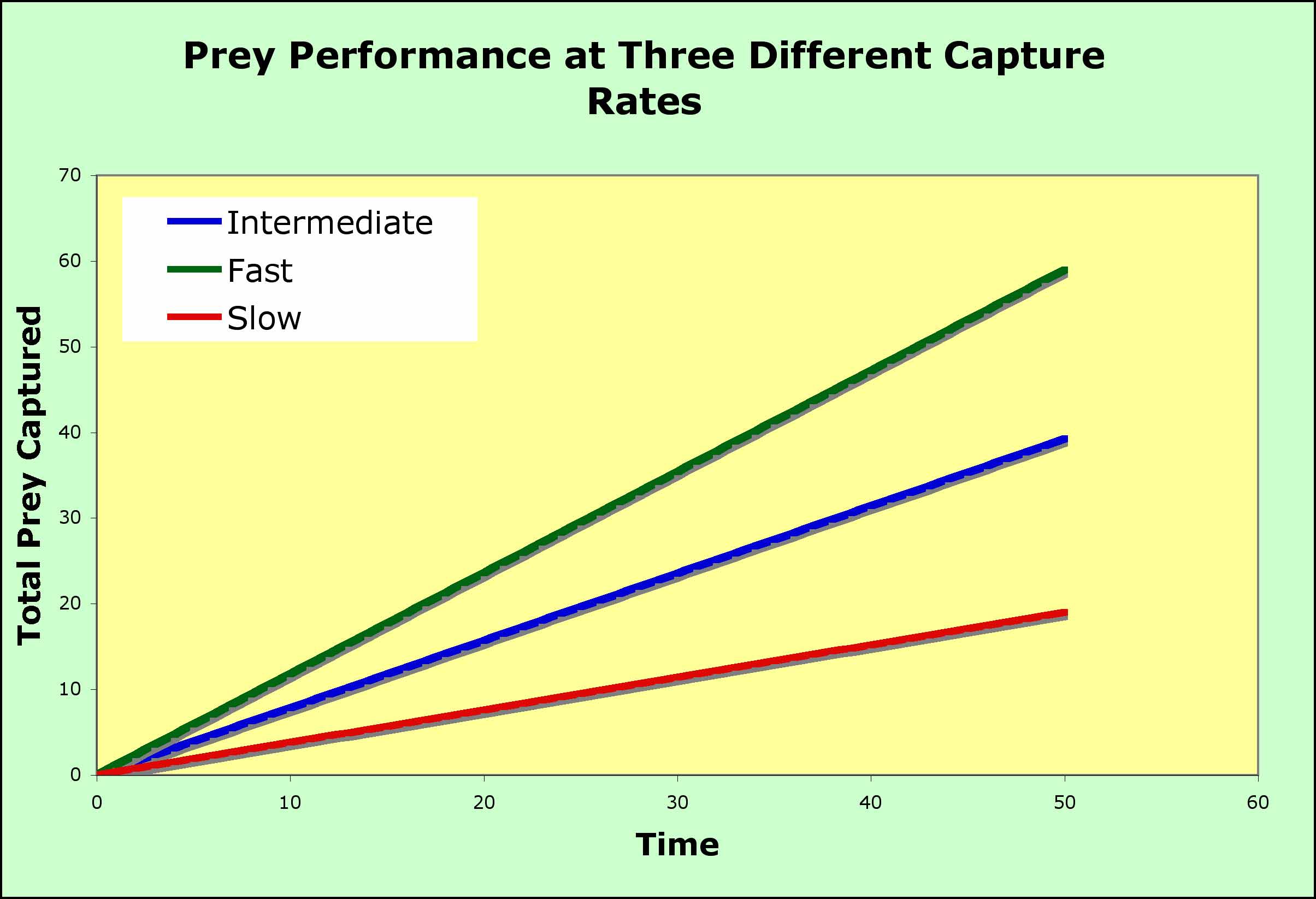
- consider the environment without patches (the graph above), where food is distributed randomly throughout the habitat
- the animal enters the habitat and begins to search. food and encounters the food randomly
- the longer the animal searches, the larger is the total catch and the increase can be predicted by a straight line
- the slope changes with the density of the prey, so that the rate of capture can be fast or slow
- the straight lines above represent the best that a predator can do in terms of prey capture given a particular density of prey
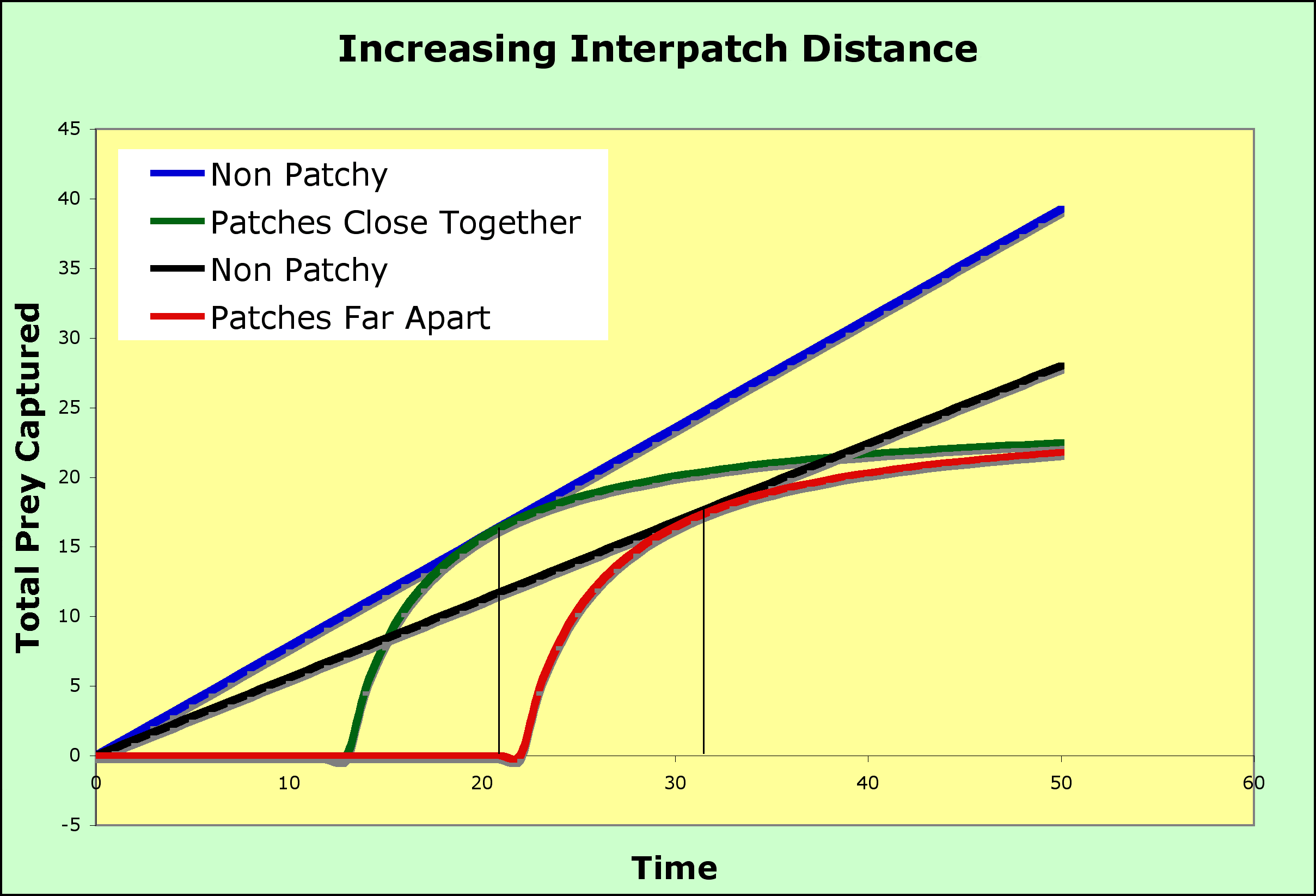
- So, what happens when we introduce patchiness into the habitat so that the prey are not randomly distributed but are found in clumps?
- The model assumes that there is a cost (in terms of time) to travel to a patch and that there will be another patch available if the current patch is abandoned, although it will cost time to find it.
- The flat portion of the green curve on the graph below, the portion that runs along the x-axis, represents the time it takes to find a patch during which no prey are captured
- After the predator arrives on a patch, it will begin to eat prey and the total intake will begin to rise.
- This rise is the upward portion of the green curve. Note that it rises very fast, which is reasonable because the predator is in a patch with prey close at hand
- At some point, the predator will begin to deplete the patch and the intake will slow.
- The slowing intake causes the total intake curve (total intake plotted versus time spent to reach the patch plus time spent in the patch) to "plateau," to flatten out, and when no new prey items can be found, the total stays flat as time goes on: no new prey, no increase in the total. Surely, it is time to leave.
- When should the predator leave the patch? before the plateau? when the plateau is reached? some time after the plateau is reached?
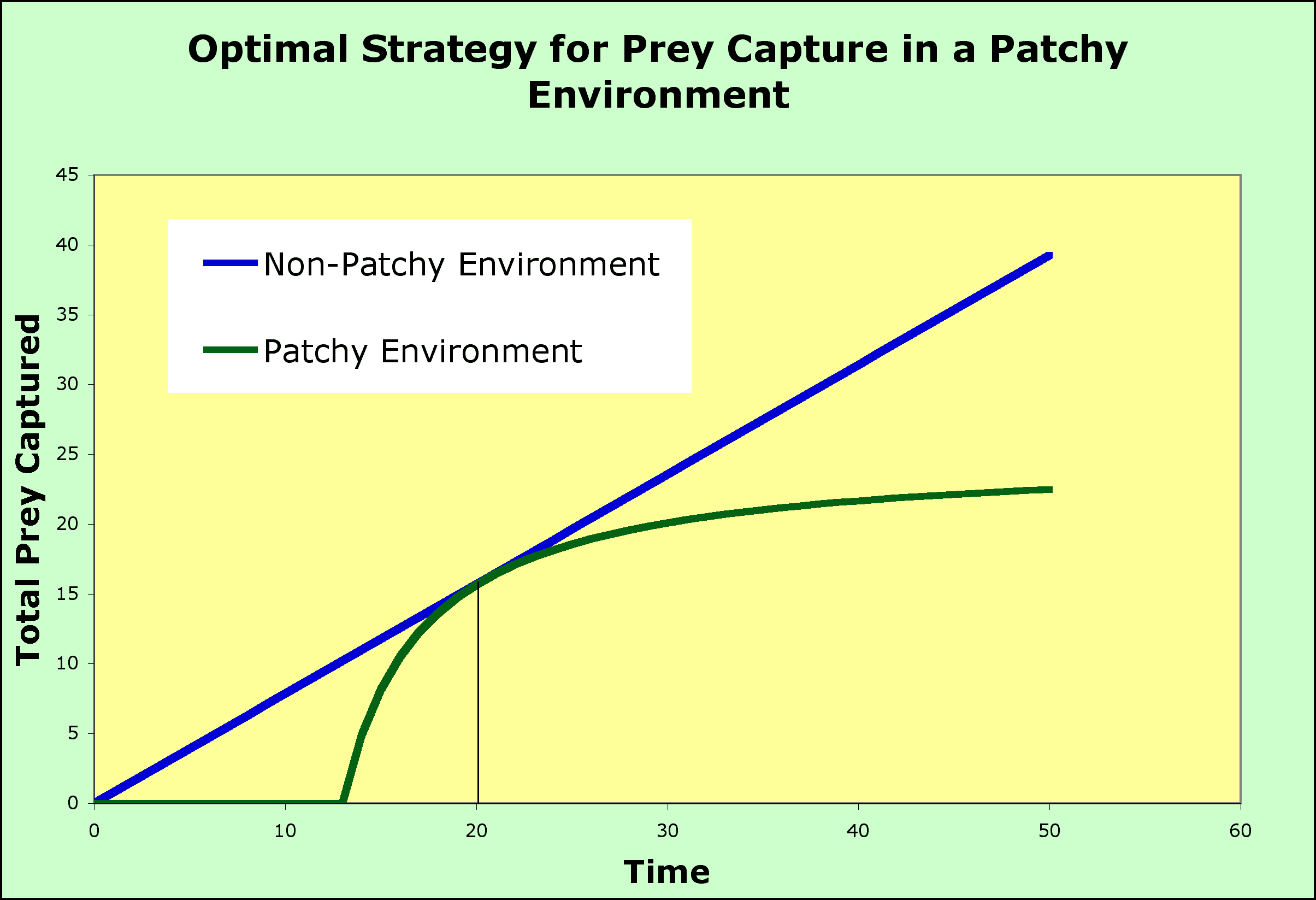
- The marginal value theorem says to leave before the plateau is reached. The predator should not wait for the plateau but should leave when the rate of return (intake of prey per unit time) is at its maximum.
- The maximum will be where the total intake in the patchy environment (the green cruve) equals the total from a non-patchy environment (the blue line) - at that point the predtor has done as well as it possibly could have but if it persists in the patch, the falling rate of prey capture means it will fall behind in total prey caught (if it leaves before the intersection point, it will not have acheived the maximum).
- By doing this, the patch is abandoned and the search for another patch begins so that the total intake over the entire search time is maximized.
- If the predator stayed in each patch until all prey were captured, it would visit fewer patches and much of its time on the patches would be spent searching for scarce prey (prey made scarce by the predator itself).
Searching time and patch size or quality both affect the time spent in a patch.
- As search time gets larger, the time spent in a patch will increase. The relative benefit of leaving is reduced due to the increased time before the next patch is found.
- In the graph above, longer search times to find a patch results in the tangent point moving to the right on the red curve where patches are far apart and the search time is longer (moved to the right compared to the tangent point on the blue curve, where patches are close together and so the search time is shorter)
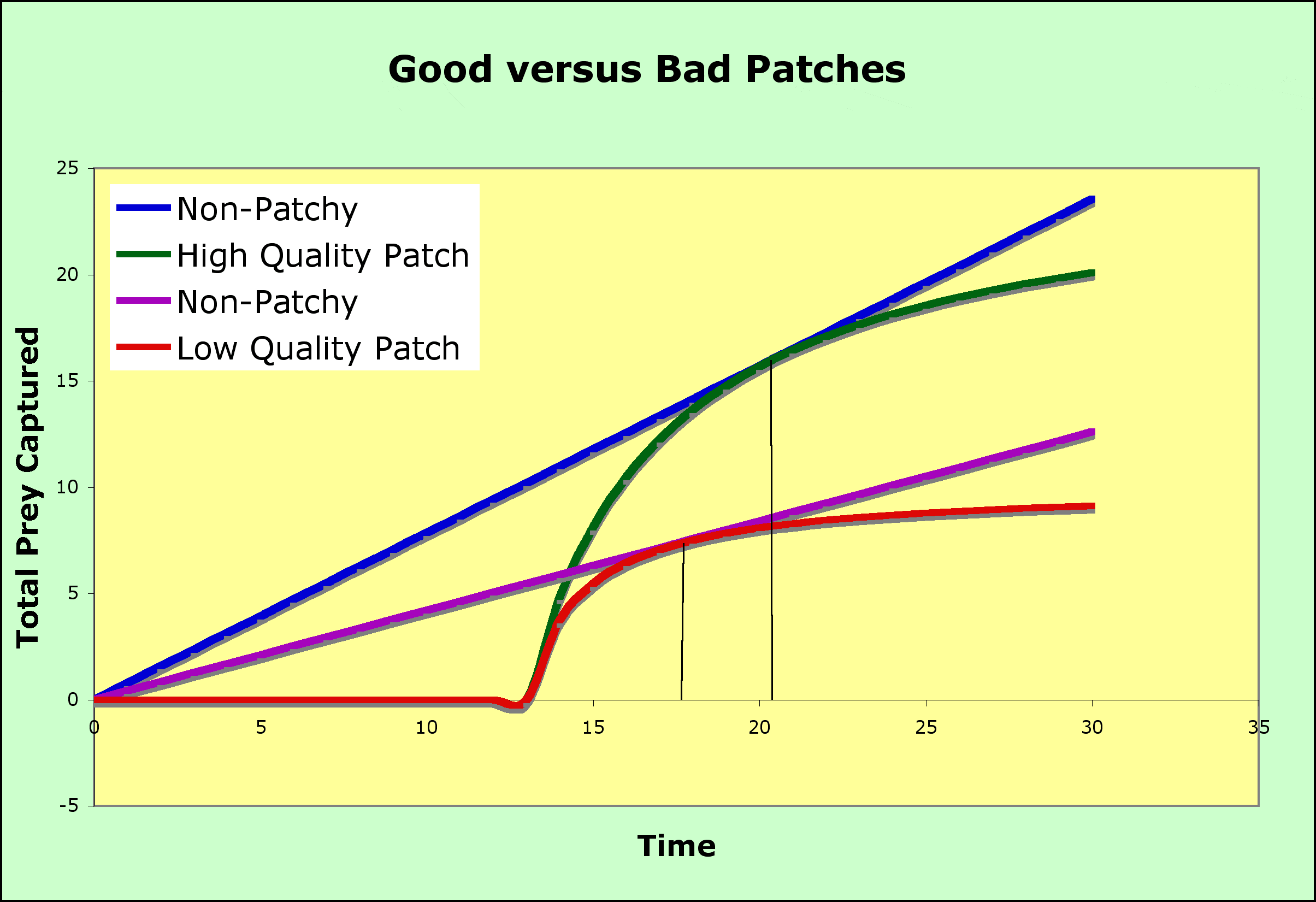
- Low-quality patches increase the time spent in a patch compared to high-quality patches
- for patches of higher quality, the prey captured curve goes up faster and plateaus higher (the green curve) than for low quality patches (the red curve)
Coevolution between Predator and Prey
Changes in the predator and prey due to either the necessity of catching prey or the risk of being eaten may mean that the one species changes as in response to the other - this is and example of Coevolution.
How important is predation in nature?
Strategies for predator avoidance
Coloration:
- This is done so that the phenotype to be avoided is reinforced in the mind of the predator
- one could argue that this is a kind of mutualism among the Mullerian mimics
Behaviors:
Polymorphism: the presence of more than one morph in the population. Each morph has to be at a higher frequency than would be produced by mutation alone
Chemical defenses in prey either make the prey too toxic or smelly or too distasteful to eat
Masting - the production of lots of young in some years, few in others
Coevolution among Herbivores and Plants
Plants cannot run from herbivores but they can try to hide (see strategies below) and they can defend themselves once attacked. This defense can take many forms and pose a problem for the herbivore. If it does not respond to a plant's defense, it must drop that plant from its diet. Thus, coevolution between plant and herbivore is expected. We will discuss the forms of defense the plant might exhibit below. As a counterpoint and to demonstrate that coevolution is occurring, we discuss herbivore responses below. However, here it might be appropriate to ask which side is winning the war? Probably neither, although the Red Queen hypothesis is probably functioning. Why neither? Two observations: We know of no plant, no matter how well hidden or toxic, that does not have at least one herbivore adapted to feed on it. We can also see that, although herbivores are legion and many leaves exhibit some evidence of herbivory by late summer, the natural world it full of plants and leaves.
Secondary Chemicals - those that are synthesized by specialized pathways in plant cells, not part of general plant cell metabolism
Two general types of secondary chemicals (based on their effect on the herbivores, not on the structure of the chemicals)
Quantitative defense
Qualitative defenses
Mechanical defenses
Failure to attract
Reproductive inhibition
Anti-herbivore mutualisms
Below-ground storage
Induced Defense versus Constitutive defense
Rosenzweig, M. L. and R. H. MacArthur. 1963. Graphical representation and stability conditions of predator-prey interactions. American Naturalist 97:209-223
Last updated February 19, 2007