| |
BIOL
4120
Principles of Ecology
Phil Ganter
320 Harned
Hall
963-5782
|
The flowers
on this cactus are not cactus flowers. They are the flowers
of a holoparasite, a mistletoe, that lives inside of the cactus. |
Lecture 15 Symbioses: Mutualism, Commensalism &
Parasitism
Email me
Back to:
Overview - Link
to Course
Objectives
Symbiosis,
Parasitism and Mutualism
Parasitism
- Parasitism is one of the +,-
species interactions
- There is no easy definition that
will separate everything biologists consider parasites from herbivores or
predators
- almost never kill the host
directly (although some diseases do this, of course)
- usually live in intimate
contact with their hosts (although some insects commonly considered parasites,
like ticks and mosquitoes, spend much of their life span not in contact
with a host)
- tend to be much smaller
than their hosts so that one host often supports many parasites
- Host
is the organism from which the parasite or parasitoid derives its sustenance
- some parasites have only
one species as a host
- many diseases infect
only one or a couple of host species
- some parasites can have many
(usually related) species as hosts
- ticks and mosquitoes
will bite any warm-blooded animal they find
- Host
Range is the number of host species one parasite species
will attack (thought to be narrower for parasites that are totally dependent
on the host for all feeding)
- some parasites require more
than one host species before they can complete their life cycle
- these parasites (many
parasitic worms [Platyhelminthes and Nematoda]) have complex
life cycles
- often two different species
as hosts (rarely three)
- often the hosts are not closely related
for parasites with complex life cycles
- schistosomiasis nematode must infect
both a freshwater mollusk and a vertebrate to complete the life
cycle
- host in which meiosis
takes place is called the Definitive Host
- other hosts are referred
to as Intermediate Hosts
- parasite reproduces asexually in
intermediate host
- Vectors
are organisms that are necessary to transmit the disease
- some vectors are not
affected by parasite and can't be considered hosts, just vehicles
to transport the parasite
- some vectors are also
hosts
- Reservoirs
are alternative hosts where the infection may remain if it is eliminated
from another population (deer are reservoirs of eastern equine encephalitis
for human populations)
- Parasites can be divided into:
- Ectoparasites
that remain outside of the host's body
- Endoparasites
that enter the host's body
- Holoparasites
(used for plants only) plants that parasitize other plants and
no longer photosynthesize but get all water and food from the host (ex:
Dodder, Dutchman's Pipes)
- Hemiparasites
(used for plants only) plants that parasitize other plants for
water and minerals, but photosynthesize to make their own food (ex: Mistletoe)
- Don't confuse hemiparasites
with epiphytes, plants
that grow on other plants but do not invade their tissues to steal
water and nutrients (ex. many orchids
- Effect of Parasite on host
- Parasites may kill (as when
a disease kills its host)
- Parasites may reduce host
fitness through lost growth or lost reproduction due to stress from harboring
parasite
- Parasites may sterilize the
host
- Parasites may alter the hosts
phenotype
- some parasites change
the sex of the host
- some parasites alter
behavior of the host so that the host acts to benefit the parasite
(at its own expense)
- Parasites
- Come from almost all taxonomic
groups
- parasitic bacteria. plants,
fungi, protists, and animals
- Attack all kingdoms, including
bacteria (which have viruses)
Mutualism
- Relationship between two organisms
that benefits both
- mutualisms carry both costs
to each partner and benefits as well
- mutualisms are favored when
the benefits are greater than the costs, so it is the net benefits (or
benefit cost ratio) that determine the outcome of these interactions
- Mutualisms can be:
- Obligatory
- organisms cannot survive in the absence of the other partner
- Facultative
- organism can lead an independent existence
- mutualistic relationship does
not have to be symmetric
- one organism may be obligated
to the mutualism, while the other can live without its mutualistic partner
- Example of asymmetry -
- many stony corals do not feed at
a rate to sustain themselves when they lose their algal partners
- the algal partners can usually grow
and reproduce outside of the corals
- Importance
of Mutualisms
- Mutualism once thought to
be among the important interactions among species
- Allee and the terrestrial
isopods
- Showed that terrestrial isopods (pillbugs
or rolly-pollys), which are very susceptible to desiccation, survived
longer in groups than when alone when the soil got dry
- Allee effect
is still used to indicate an situation in which animals are better
able to survive and reproduce in groups than when alone
- interpreted this to mean that organisms
often cooperated for mutual benefit
- makes cooperation as important as
competition and predation (more negative interactions)
- Mutualism fell out of favor:
- Competition/predation
studies became more common
- Theory predicted that
conditions that favored mutualisms were narrow and unlikely to be
found in nature.
- Current view has restored
mutualisms as important in natural systems
- important mutualisms
identified (mycorrhizae, nodulation, arthropod-bacterial nutritional
mutualisms, etc.)
- theory modified to widen
the conditions under which mutualism is favored by selection.
Relationship between parasitism
and mutualism
- Many parasitic relationships
evolve to reduce the cost for the host
- benefits the host in that
the parasites do less damage
- benefits the parasite in
that there will be more hosts to parasitize if they are not excessively
harmed by the parasite
- If the process crosses the line
where the benefits outweigh the costs, a parasitism can become a mutualism
- This model of the origin of mutualisms
solves the "origin problem."
- How do mutualisms begin if
the benefits only arise when the partners come together? Were the
adaptations serendipitously already there?
- This seems not likely
due to the prevalence of mutualisms.
- Did the first insects to visit plants intend
to be pollinators? Probably not, they were probably just foraging.
- Plants that attracted the insects to
their reproductive structures benefited from the accidental pollinations
- From the accidental beginning, where
the plants were mostly harmed by the insects, the elaborate pollination
systems (including flowers, floral rewards, specialized animal
behaviors, etc.) developed over time to increase the benefits
of the interaction to both partners.
- Probably not the origin of all
mutualisms
Symbiosis
- a relationship between individuals
of two different species in which individuals of one species lives on or in
individuals of the other species
- mutualisms may or may not be
symbiotic
- lichen fungi and lichen algae
are only found together - symbiotic
- plants and pollinators are
only in contact when the pollinator is feeding - not symbiotic
- parasitic interactions may or
may not be symbiotic
- parasitic tapeworms can only
grow and reproduce in the gut of a vertebrate and only leave one host
to get to another - symbiotic
- mosquitoes spend as little
time on their hosts as possible (for obvious reasons) - not symbiotic
Diffuse
Mutualisms
- these are called diffuse because
the strength of the connection between any two species is not as strong as
when each species has one and only one other species with which it can form
a mutualism
- may involve many species
- yeast living together often
have mutualisms in which each can feed from the activity of the others
present and the interaction may involve three or more species
- each partner may interact with
more than one other partner
- the species involved in the
mutualism may change from place to place or through time
- some plants have many species
as pollinators, including birds, bats, and insects
Modeling
Mutualism
We can modify the logistic equation
to model mutualism, just as we did for competition. The difference this time
is that we assume that the presence of a mutualist has the opposite effect that
the presence of a competitor did. A mutualist will increase the carrying capacity
of the environment, and the size of the effect will increase as the number of
mutualists does.
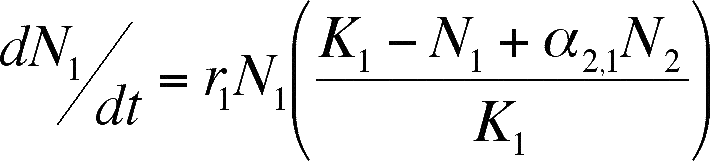 and
and 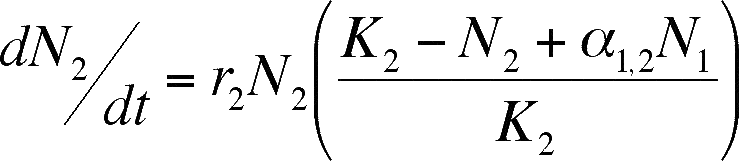
- The equations above do this (notice
how close to the Lotka-Volterra equations they are). Each equation is identical
to the Lotka-Volterra equation but the sign of the alpha term has been changed
to positive. Thus, addition of the mutualist species adds to the total number
of individuals that can be sustained in the population (K is supplemented,
not reduced as it is in competition).
- These equations can be solved
to get a zero-isocline and analyzed in the same fashion as in the competition
model.
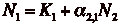 and
and 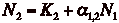
- Now the lines have a positive
slope, but K1 and K2 haven't changed.
- Facultative Mutualism - in the
model graphs below, both species are facultative mutualists. This means both
have a positive carrying capacity because each could exist in the environment
without the presence of the other. But the slope of the zero isoclines is
positive, so each goes to the right and up from K1 and K2.
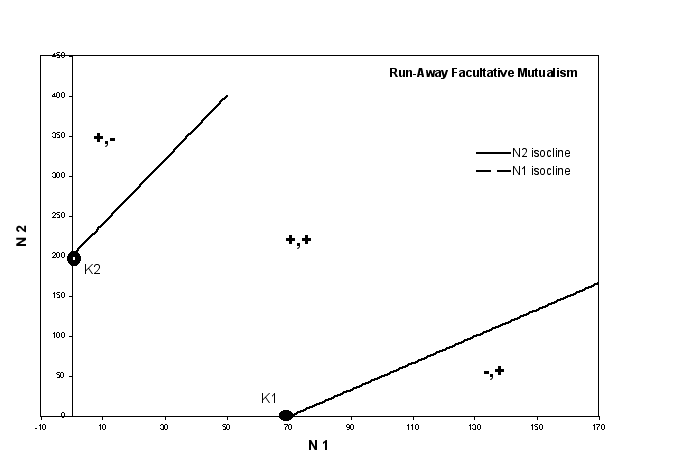
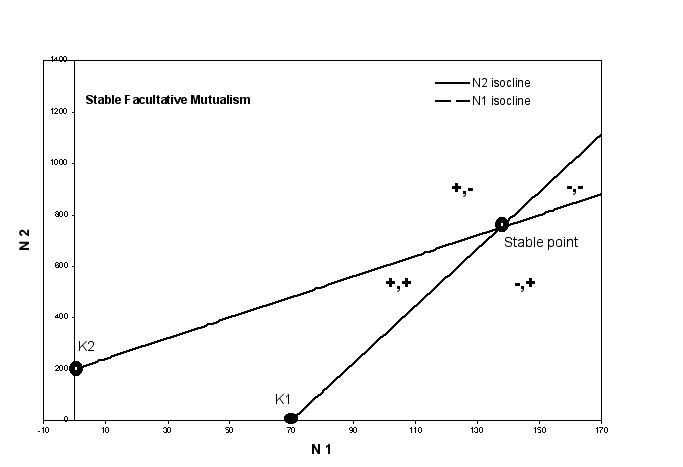
- the + and - signs indicate
the growth rate of each species when the system is in a particular region
(the first of the pair is always species 1, the second is always specie
2)
- notice that, in both graphs, the
+ region is consistent. For instance, all space to the right of K1
is negative for species 1, indicating that in this region the population
size of species 1 is too great, it is beyond the carrying capacity,
even accounting for the presence of he other species (which increases
the populations size in this case).
- If you go to the notes for
the chapter on competition, you will see that the + and - regions
are consistent between these graphs and the graphs generated by
the Lotka-Volterra equations
- Now to the key point. What conclusions
can we draw from each graph.
- The Run Away graph on top
describes a case in which the zero isoclines do not cross. This
means there is no point at which both species are just maintaining
their population sizes. In other words, there is no equilibrium
point with both species present. What happens, if you follow the
vector changes around (see the book), is that, no matter where
you begin in this system, you end up with a run-away situation,
in which one species always increases the size of another species,
which, in turn, increases the size of the first, which increases
the size of the second . . . until both species hit infinite population
sizes.
- The Stable graph on the bottom
has an intersection point. where both species' growth rates are
0, so that it is an equilibrium point. If you follow a point around,
you will see that it is a stable equilibrium point and no matter
where you begin, you always end up at the stable point. Thus,
we can see that a stable mutualism is possible even with these
simple equations.
- Obligate Mutualisms
- in the graphs below, both species are obligate mutualists. This means that
neither can exist in the environment unless the other species is present at
some critical population size. In this case, the obligate nature of the relationship
can be seen in the fact that the carrying capacity for each species is below
0. It is hard to interpret a negative carrying capacity, but I suppose one
might take the magnitude of K as a measure of just how unfavorable the environment
is.
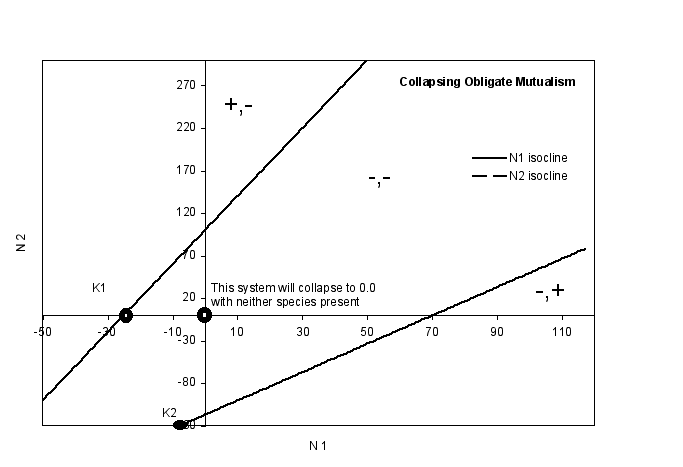
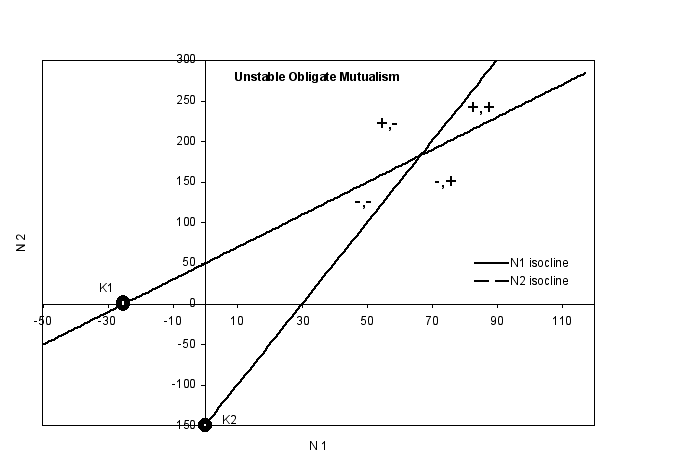
- In the Collapsing graph on
top, there is no equilibrium point with both species present. If you use
the + and - signs to follow a point through time, you will see that, no
matter where you start you end up at the origin
- this means that neither species can
exist in this environment, no matter how many of the mutualists are
present.
- In the Unstable graph on
the bottom, there is an equilibrium point, where both species can exist
in the environment even though neither could be there alone. If you follow
a point around, you will see that you never go back to the intersection
of the lines, so that this in an unstable equilibrium
- this means that this obligate mutualism
is unstable, and any change in the population size of either member
of the mutualism will mean the collapse of both populations.
- The finding of greater instability
for obligate mutualisms may be an outcome of the model that is not well supported
in nature, as many obligate mutualisms exist
- Note that the book has some
more sophisticated models, in which the zero isoclines are not straight
line but are curved, and that this opens up the possibility of stable
obligate mutualisms
- this might mean that nature
is more complex than our simple models, but I think we should at least
explore how things might interact with the simple models before going
on to more complicated models.
Modeling
Parasitism
Epidemiology
is the science that studies disease
- Infectious diseases are caused
by parasites and are the most intensively studies parasite systems (due to
their importance to our health)
- parasites are referred to as
pathogens in epidemiology
Factors in the spread of disease
Rp = SBL
Some consequences:
- The longer the host is infective,
the greater the replacement rate of parasitized hosts, so there is pressure
on the parasites to keep the host alive (increase L, increase Rp)
- High transmission rates (large
B) leads to greater replacement rate of parasitized hosts, so there is pressure
on the parasites to evolve greater rates of transmission (increase L, increase
Rp)
- Given the limited resources
of the host, it may not be possible to do both of the above
An important consideration (for
the parasite) is NT
NT
= threshold population
of susceptible hosts at which Rp = 1
and below which Rp <1
If Rp in the first
equation is set to 1, then
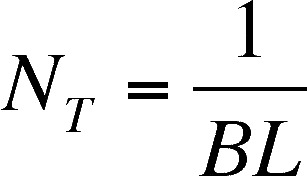
Some consequences:
- If NT is not constant,
an increase in either transmission rate or infectious period will reduce
the size of the host population needed to maintain the parasite
- If NT is constant,
then an increase in one parameter (either transmission rate or infectious
period) will lead to a decrease in the other parameter (in other words,
an increase in transmission rate will reduce the infectious period and vice
versa)
Epidemiologists often try to define
NT so that they can predict the critical
density of a susceptible population
Evolution
and Mutualism/Parasitism
- Evolution of parasitism and mutualism
are excellent examples of the process of Coevolution
- Coevolution
is the process of evolutionary change in two species in which each
changes in response to change in the other species
- Coadaptation
is a characteristic of an organisms that is involved in the mutualism/parasitism
by interacting with some feature of the other partner
- an example is the communication
that goes on between roots and nodulating bacteria
- Coadaptations need not be
the product of coevolution
- Serendipity
- good fortune due to chance - can also bring together two organisms
that already have features that make their mutualism possible
- Conflict within Mutualisms
- Stable mutualisms must prevent
cheating by a partner (getting benefit, bearing no cost)
- Parasites and Hosts are also
coadapted
- Coadaptation
often due to arms race type of coevolutionary
changes in host and parasite
- Parasites differ with respect
to their host specialization
- Monophagous
parasites attach a single species of host
- Polyphagous
parasites attack several species of hosts (usually they are related)
- Endoparasites are, in general,
more often monophagous than are ectoparasites, although there are many
exceptions to this observation.
The
Impact of Parasites:
- Host defenses
- Cellular Defense
Reactions
- Encapsulation of parasite's
cells (often reproductive cells) by the host so that they are non-functional
- Cell surface changes
- Change the marker
molecule and the parasite may not recognize the host
- Immune response
- Grooming and preening
to remove ectoparasites
- Epidemics
- Often can see the effect
of an addition of the parasite to the host population as an epidemic (outbreak)
of a disease
- Difficult to remove the parasite
from a natural population and so it can be difficult to do field experiments
with parasitic systems
- If this were not so,
we would have performed many such removals in trying to cure us
and our crops and livestock of disease
- Disease may show cycles similar
to predator-prey cycles (in humans, whooping cough and measles show this cycling)
- basis is the proportion of
susceptible hosts
- susceptible hosts become non-susceptibles
after infection, as immunity's memory system makes a second infection
unlikely
- After an outbreak, enough hosts become
immune to drop Rp below 1, so the disease declines in the population
- As disease prevalence falls, new
individuals entering the population (births and migration from populations
without the parasite) boost the proportion of susceptibles
- When this proportion is high enough to
boost Rp over 1, another outbreak begins, starting the cycle over
again
- Can you see why this cycling is most
apparent in diseased that affect children?
- Disease can set limits to the
population size or the distribution of a host or hosts
- Rinderpest
in Southern Africa - virus with wide
host range (large, grazing mammals)
- Buildup of host (cattle) after establishment
of European-style ranching
- Outbreak of parasite
after introduction of diseased cattle from Southeast Asia caused decline
in cattle
- also led to loss of natural
populations of other hosts
- Decline in wild populations of
large grazing animals (antelopes, gnu, etc.) lead to:
- change in vegetation over
wide areas
- reduction
of tsetse fly population, which feed on large mammals
- Decline in tsetse fly population
lead to decline in cases of sleeping sickness caused by a trypanosome
transmitted by the tsetse fly from human to human and from other large
mammals to humans
- Competition can be mediated through
parasite - called Apparent
Competition
- White-tail deer and Parelaphostrongylus
tenuis
- White-tail deer are tolerant
- Other cervids (moose, other
deer like the mule deer, pronghorn) are harmed
- Where white-tail act as a
reservoir, other cervids do not occur
- Evolution may change the character
of host and parasite
- Evolution of resistance to antibiotics an
example of the evolutionary potential of parasites
- Virulence (transmission rate
and infectious period) may vary through time
- Rabbits and Myxomatosis
- Less virulent strain of virus evolved
- When both are present
in a rabbit, virulent strain grows faster, overgrowing the less virulent
strain, and wins by being the strain transmitted to the next host
- When alone, less virulent strain
meant that rabbits would live longer, infect more bloodsucking
insects (vector)
- More vector meant the
less virulent strains had a higher rate of transmission as the rabbits
lived longer to be bitten and the less virulent strains would win
at the global level, although it loses at the individual rabbit level
to more virulent strains
- after time, Myxomatosis
became a non-lethal disease and now a second virus, Calcivirus,
is being used
- Biological
control through the use of parasites
and parasitoids:
- Attempt to reduce
the population of a pest to an acceptable level through manipulation of
the population ecology of that pest
- Note that it says reduction
and not elimination of the pest
- elimination may sometimes
occur buy the usual outcome is the reduction of host to lower population
levels than without the parasite
- Not all biological control
involves parasitism
- Herbivores and predators
are also used
- sterile male release
also used (screw worm)
- Strategies can attack either
death rates or birth rate (or both)
- Death rate strategies
- Rabbits and Myxomatosis
- Poses potential problems
as the disease might jump to new hosts in the new environment
and kill non-target species
- Birth rate programs
- Sterile male programs
- Med fly and Screw
worm programs
Examples
of Mutualisms
Pollination
- Pollinator may get:
- Food (nectar, pollen- high
energy or high protein food)
- Mating advantage - some bees
get scent molecules
- Nesting materials - some
bees get wax for their nests
- Flowering plant gets:
- Efficiency of pollen transfer
(compared to wind)
- Mixing of pollen from many
plants and prevention of inbreeding
- Pollinators include flies, bees,
wasps, bats, beetles, birds
- any animal that visits the
flower regularly may be a pollinator
May be a very "tight",
highly coevolved relationship or a diffuse relationship
- Examples of diffuse systems
- Many flowers in the fields
in Tennessee are visited by more that a dozen species of insect, all of
which may act as pollinators (I have seen 10+ species of insect visiting
a flowering fruit tree at the same time)
- Examples of highly coevolved
systems
- Orchids and pollinators
- many orchids are pollinated by a
single species of insect
- flowers of orchids are often shaped
so that only the correct insect can get to the nectar and so will
carry the pollen
- Fig- wasp
- there are many species of fig - they
produce many flowers enclosed in a capsule (we call the capsule and
its contents a fig)
- each has its own species of wasp
(called Agaonid wasps)
- the female wasp lives all of its
larval life in fig and only spends enough time out of one as an adult
to disperse to the next fig, where she will deposit her eggs and never
leave (only its progeny will)
- males never leave the fig in which
they hatched, grew as larvae, and pupated
- fertilize females in same fig and
die there, never having left it
- the fig must supply food for its
wasps or it will not produce a new generation
- wasps must not overexploit the resource
or they will eat the fig and it will never produce the next generation
of fig plants
- neither species can enter a new environment
without the other
- Yucca - moth
- similar to fig story - each species
of yucca is pollinated by a single specie of moth which lives only
on the species of yucca that it pollinates
Some plants and some animals cheat
- some animals may take nectar
but do not carry pollen
- some insects are unable to
get to the bottom of deep, vase-like flowers but simply drill through
the base of the flower to steal nectar
- some plants look just like other, nectar-producing
flowers, and so trick the pollinator into visiting them without the cost of
rewarding it
- some plants have flowers that look and smell
like females of insects. They attract the males, who mate with the flower
and carry away pollen
Dispersal
Mutualisms
- Fruits are plant rewards for
animal dispersal of seeds
- Seeds often pass through the
guts of dispersers without harm
- some seeds even benefit from
this by being deposited with the manure as a fertilizer
- some seeds use the passage
as a signal to germinate and will not do so without this
- some plants protect the seed
with toxins while making the fruit palatable
- peach seeds (pits) are full of cyanide
- some plants sacrifice some
seeds to dispersers (seeds are usually very good food - lots of vitamins,
protein and lipids)
- Lots of cheaters in this system
(whenever seeds are eaten as food and are not just passing through the gut)
- Fruit colors are important signals
- make fruit apparent to dispersers
(advertisements)
- green fruit often contain
same toxins as other part of plant to stop herbivory
- when ripe, color change signals readiness
in that the fruit has:
- lost it toxins
- been stocked with sugars
Cleaning
Mutualisms
- one species gets food by removing
(and eating) ectoparasites of another
- partner loses its parasites without
having to clean itself
- happens on reefs where cleaner
shrimp clean parasites from fish at "cleaning stations"
- also on reefs, cleaner fish
perform same function as shrimp
- oxpecker birds eat parasites
from outside of large herbivores (cattle, antelope, rhinoceros)
- although they keep the ticks, etc.
off, this may not be a mutualism, as the oxpecker will peck a vulnerable
area (often an ear) and drink blood when parasites are not available
Defense
Mutualisms
- one species gets food and/or
shelter from another species
- other partner gets protection
from being eaten
- Ant-Acacia system
- Bull Thorn Acacia provides:
- place for ants (Pseudomyrmex)
to live in swollen base of acacia thorns (hence the name bull-thorn)
- food for ants in form of
special extension of leaves call Beltsian
bodies
- ants are aggressive and attack almost
anything that comes into provide protection from
- other insect herbivores
- large, vertebrate herbivores
(including you, if you happen to lean on the tree)
- Some defensive mutualisms involve
plants and fungi
- Some grasses are infected
with fungi (Clavicepts and other Ascomycetes) - long though to be parasitic
but the fungi are the source of alkaloids
- the alkaloids are protection
from herbivory as they are toxic and bitter
- some evidence that infected
plants grow faster and produce more seed.
Bacteria
- Aphid, Leaf Hopper Mutualism
- Aphids and leaf hoppers feed
on sugary sap sucked directly from the phloem tubes of plants
- sap is a poor diet that is
high in sugars, low in amino acids
- insects have essential amino
acids, just like us, and so they cannot live on this diet without help
- Bacteria live inside special
cells called Bacteriocytes in
the fat bodies of the aphid and leaf hopper
- Bacteria receive sugars from
plant via the aphid and supply the aphid with amino acids
- Bacteria also receive easily-made amino acids
from insect and transform them into essential amino acids that the insect
cannot make
- Without bacterial mutualists,
aphids and leaf hoppers could not live as they do, so this is an obligate
mutualism for the insects
- Bacteria are adapted for only
one environment, inside insect cells, and so they are also obligate mutualists
but they might have begun the relationship as parasites
- All species of aphid and leaf
hopper have a nutritional mutualism, usually with bacteria but a few species
of each have fungal mutualist partners instead of bacteria
- In one case, the leaf hopper
does not excrete the uric acid it produces as its nitrogenous waste but
recycles it to the yeast, which use it in amino acid synthesis
- In the same insect, the yeast
synthesize proteins that are stored in the eggs and, without these proteins,
the eggs will not produce viable zygotes when fertilized
- This sort of mutualism, in which
specialized cells harbor microbial mutualists that are needed to improve the
quality of the animal's food, are not confined to aphids and plant-hoppers
- they also occur in some beetles
(Anobiid Beetles) that live in dead wood (called powder-post beetles because
of the wood dust from their boring activities)
- Some insects called Lacewings
(Neuroptera) are important in biological control because their larvae
eat aphids and planthoppers but the adults of some species feed on plant
sap and have yeast in their guts that provide them with required
amino acids and lipids
Lichens
- Many fungi are lichenized, each
one needs a particular species of algae
- each algae species usually
can form a lichen with several different species of fungi
- because the fungus is the
unique partner in each lichen, it is the fungal name that becomes the
lichen's name
- Fungi get photosynthate from
algae
- algae get minerals and some desiccation
protection and dispersal from fungi
- Obligate mutualism
Plant
- Mycorrhizae (and some bacteria)
- Very common and very important
mutualism - these fungi can be 50% of the
microbial biomass in soils
- two important types:
- Ectomycorrhizae
- many species of both Ascomycota (ascus-forming fungi) and Basidiomycota
(club-spored fungi), the two largest fungal groups - many common mushrooms
are the reproductive structures of ectomycorrhizal fungi
- wrap hyphae around roots, do not
penetrate cell walls of plant cells
- hosts are trees (many
conifers) in temperate or boreal systems
- Vesicular-Arbuscular
Mycorrhizae (VAM, sometimes the Vesicular part is
dropped and they are called just AM) - come from a few genera of Zygomycota
(the group bread molds belong to)
- hyphae have no walls (septae), so
the entire mycelium (all the thread-like hyphae) are essentially
a single cell (this condition is called coenocytic).
- hyphae penetrate the cell walls and
split into lots of bifurcations that end in vesicles (swollen
tips), but the hyphae do not penetrate the cell membrane, which
folds inward to accommodate the fungal growth
- almost any plant that does not have
an ectomycorrhizal association will have a VAM association (the
majority of plants by far)
- have BLO's inside their hyphae -
first called Bacteria-like organelles, now known to be intercellular
bacteria - role in the system not known at this time
- Plants get several benefits -
- minerals from absorptive
power of fungi
- hyphae of fungi increase
the absorptive area of roots by penetrating the soil much more finely
than the roots can
- growth rate and reproduction
of plants often much lower if mycorrhizae are removed
- protection from pathogens,
both bacterial and fungal
- some plants even get
their carbohydrates from mycorrhizae (see orchid section below)
- Fungi get photosynthate from
plant
- Facultative mutualism, except
for orchids
- Orchids and Orchid
Mycorrhizae
- all Orchids have important pollinator mutualisms
with insects (see above) and also important fungal mutualisms
- orchid seeds are tiny and have little stored
resources (fats, carbohydrates, proteins) for the germinating embryo
- Orchid mycorrhizae in soil (or on surface
of a plant for epiphytic orchids) penetrate the seed coat and trigger
germination of the seed, then supply the young plant with sugars and proteins
until it becomes photosynthetic and can return the favor
- some orchids are non-photosynthetic and the
mycorrhizae continue to supply sugars and proteins that they get by penetrating
the plants the orchid is growing on - in this case the fungus is a parasite
of one plant (the tree) and a mutualist of another (the orchid) at the
same time!!!!!
Plants
- Nitrogen-Fixing Bacteria
- Nitrogen is a form useable by
plants (nitrate, nitrite, or ammonium) is the product of the metabolism of
other organisms
- N2 is plentiful
in atmosphere but useless to plants
- the process of making N2
into organic nitrogen (as the above forms of N are collectively called)
takes lots of energy
- bacteria (Azotobacter, Azobacter,
some Pseudomonas species, some blue-green algal species) are free-living
microbes that can fix nitrogen
- the bacteria are all anaerobes
and live in regions of the soil where oxygen has been depleted (see
section on coadaptation and leghemoglobin below for why this is so)
- Some plants have a mutualism
with bacteria to transform atmospheric nitrogen into organic nitrogen -a very
important mutualism
- Lack of nitrates (and derived
compounds) often limits plant growth in terrestrial ecosystems
- Ability to produce organic N locally is a
great advantage in nitrogen-poor soils
- many plants in the Fabaceae
(also called the Leguminosae - the pea family that includes peas, beans,
clover, alfalfa, honey locust trees, and many more trees) and other families
can nodulate
- Rhizobium is the genus of
bacteria that participate in nodulation
- Presence of bacteria causes plant
roots to nodulate
- Nodules provide bacteria
with a place to live and an environment conducive to their growth
- Plant responds to chemical
signals produced by bacteria
- secretes chemical attractants for the
bacteria, which migrate to root and enter it
- presence of the bacteria and their secretions
promotes cell proliferation by plant to make the nodule
- Plants pay a price for a ready
supply of organic N
- supply photosynthate to bacteria
for growth and for the expense of fixing N
- must also maintain the proper,
oxygen-depleted environment for fixation
- nitrogenase, the enzyme
that catalyzes the fixation, is sensitive to the presence of oxygen
- oxygen fits into
its active site as well (or even better) than does nitrogen, so
it poisons the process if it is present
- the plant and bacteria
have a coadaptation that produces the low oxygen
environment needed
- oxygen is soaked
up by the presence of a compound, Leghemoglobin,
that binds to oxygen
- Leghemoglobin
is related to our hemoglobin, both through structure and ancestry
- the protein portion
is produced by the plant from genes in its nucleus
- the heme portion
is produced by the bacterium with enzymes encoded by genes
on its chromosome
- this is a Facultative mutualism
- pea family plants all grow
without nodules (but more slowly)
- bacteria grow in soil without
pea plants (but much more slowly)
Hard
Corals - Algae
- Corals get photosynthate from
algae
- algae get minerals extracted
from sea by animals
- free-floating algae are "trapped"
in the water drop in which they float
- only get nutrients that diffuse into
their neighborhood and diffusion is a very slow process
- algae in corals are fixed
and waves pass by
- they extract nutrients from many
gallons of water each day, not just from the drop in which they are
floating
- the difference is huge
- waters surrounding reef are usually
very clear - indicating that they have little algal growth (low productivity)
- reefs are as productive as tropical
rain forests, among the most productive systems on earth
- Facultative/Obligate symbiosis
- Algae can leave when conditions
not right (bleaching of coral)
- Coral can feed by predation
on plankton (but growth is slow or even negative)
Giant
Clam - algae
- Clam gets photosynthetic output
of algae
- algae get minerals absorbed by
clam and protection from herbivores
Yeast-Drosophila
Mutualism
- Yeast need to disperse
from habitat patch to patch
- Yeast spores are not resistant to desiccation
so they must be carried
- Insects need high protein diet
- plants often low in protein,
which are needed for making eggs as adults (even when eaten by larvae)
- most yeast grow in dead plant material
- yeast are much higher in protein
than the plant tissue they eat and so are high quality food for insects
- Cactophilic yeast- Drosophila
- 10-20 species of fly, all
found only in cacti
- 20-30 species of yeast, most
found only in cacti
- mutualism is diffuse but
obligatory
- Flower Beetles and Yeast
- Flower Beetles (Nitulid beetles)
and yeast have a mutualism very similar to Drosophila and yeast
- the beetles carry yeast
from flower to flower
- the yeast use the
flower for food
- the beetles eat the
yeast
Coral-Crab
Mutualism
- Hard corals need sunlight (see the coral-algae
mutualism above)
- Overgrowth of corals by seaweeds (macroalgae)
can shade them and kill them.
- Some corals (Oculina arbuscula is
an example) avoid overgrowth because an herbivorous crab (Mithrax
forceps) forages on the algae.
- When the crab is present, no overgrowth occurs,
the coral grows faster and survival is greatly increased.
- The crab gets not only the algae as food.
It lives in the coral and avoids predation as a result. So the
coral is not just food, it is a protector.
Agricultural
Mutualisms
Ant
- Aphid Mutualism
- Aphids are protected by ants
- Ants get sweet plant sap from
aphids
- Ants are like ranchers, as they
move the aphids from place to place on the plant to take advantage of where
most sap is available
- So, considering the Ant-Aphid
mutualism and the Agricultural mutualism, it appears that we did not discover
either farming or ranching, or, if we did, we did not discover it first.
- Bees and Microbes
- by the way, not
only do insects farm and ranch, but they are bakers as well.
- Some bee species feed their larvae on pollen,
but not before it has been mixed with fungi and bacteria and allowed to
ferment (like a baker allowing the bread to rise)
- the fermented mixture is referred to
as "bee bread"
- the microbes provide nutrition and turn the
pollen, which will not support the larvae alone, into a high quality diet
- the bees transport the microbes to new nests
Commensalism
- Situations in which one species
benefits from the presence or activity of another species, but the other species
gains no benefit nor suffers any harm
- commensal organisms might
evolve into either parasites or mutualists
- Phoresy
- when one organism attaches
itself to another as a means of dispersal
- common way to disperse seeds,
animals not harmed
- small animals hitchhike
on larger animals
- Bird - Pollen mite
- when birds drink from a flower, pollen
mites (feeding on the pollen in the flower) jump on their beaks
and nestle into their nostrils
- mites jump off at next flower without
harming bird
- Burrowing animals often have
commensal organisms living in the burrows
- Can happen after the burrow
is abandoned
- many vertebrates live
in burrows made by other species
- Can happen when host is still
living in burrow
- Clams in worm burrows
on mudflats
- Clams are found no where else, so
this is obligate for them
- No evidence that the host worm benefits
or is harmed by presence of clam
Literature
Cited
Silliman, B. R. and Y. Newell. 2003.
Fungal farming in a snail. Proceedings of the National Academy of
Sciences of the USA 100:643-648
Silliman, B. R. and J. C. Zieman. 2001.
Top-down control on Spartina alterniflora production by periwinkle
grazing in a Virginia salt marsh. Ecology 82:2830-2845
Terms
Parasitism, Host, Host Range, Complex Life Cycle,
Definitive Host, Intermediate Host, Vector, Reservoir, Ectoparasite, Endoparasite,
Holoparasite, Hemiparasite, Epiphyte, Mutualism, Obligatory Mutualism, Facultative
Mutualism, Allee effect, Symbiosis, Diffuse Mutualism, Epidemiology, Pathogen,
S (Susceptible Host), B (Transmission rate), L (Infectious Period), Rp
(Replacement Rate of Infected Hosts), NT (Threshold Population
Size of Susceptible Hosts), Critical Density, Coevolution, Coadaptation, Serendipity,
Arms Race, Monophagous Parasite, Polyphagous Parasite, Host
defense, Cellular Defense Reaction, Immune
Response, Epidemic, Rinderpest, Apparent Competition, Biological Control,
Pollination, Cheating, Dispersal Mutualism, Cleaning Mutualism, Defense Mutualism,
Beltsian Body, Bacterocyte, Lichen, Mycorrhizae, Ectomycorrhizae, Vesicular-Arbuscular
Mycorrhizae, Orchid Mycorrhizae, Nitrogen-Fixing Bacteria, Leghemoglobin,
Ambrosia Beetle, Commensalism,
Phoresy
Last updated February 26, 2007
