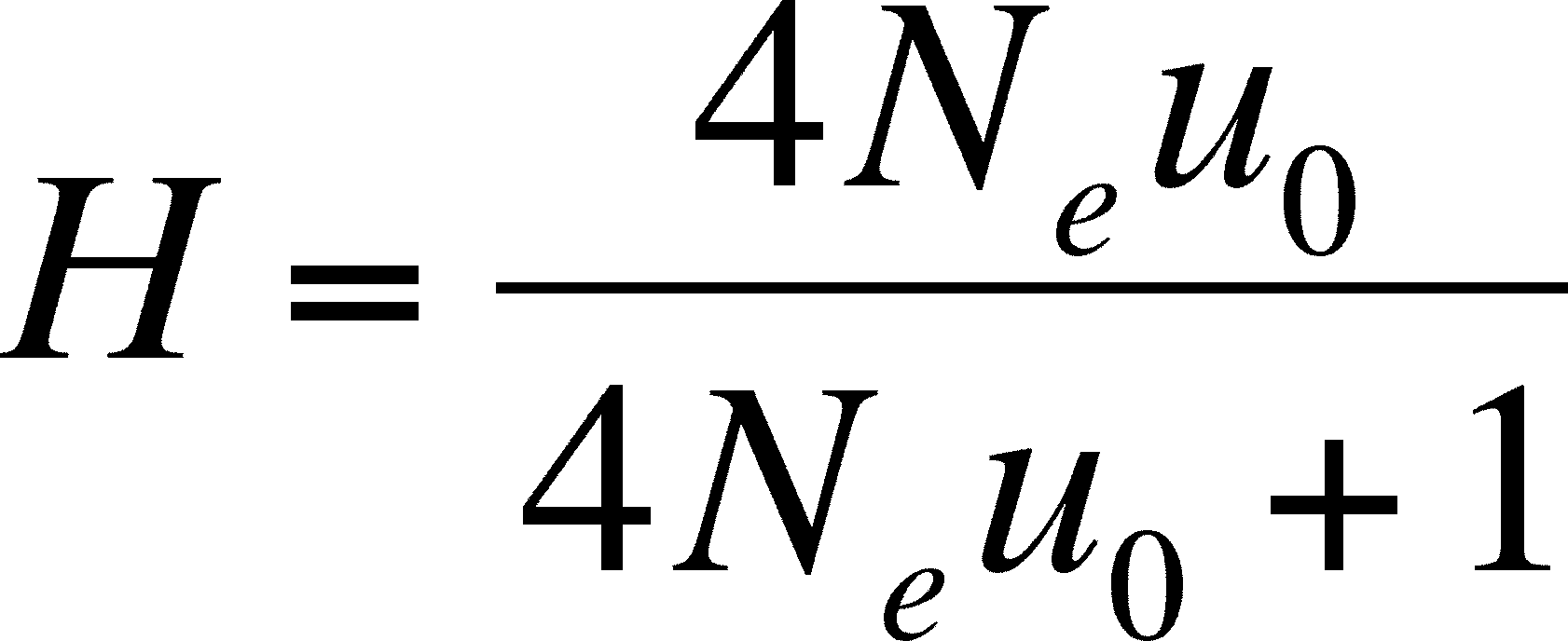|
BIOL 4160 Evolution Phil Ganter 301 Harned Hall 963-5782 |
Does the fringe on the palm trunks above have a function? |
|
|
BIOL 4160 Evolution Phil Ganter 301 Harned Hall 963-5782 |
Does the fringe on the palm trunks above have a function? |
|
Genetic Drift as an Evolutionary Force
Link to a list of Specific Objectives for lectures
Back to:
Academic
Page |
Tennessee
State Home page |
BIOL4160 Page |
Ganter
home page |
Random (chance) influence versus Deterministic influence
- Inevitability of random influence
- Genes in any generation are a sample of those in the previous generation and all sampling is subject to sampling error - deviations from expectation due to chance
- Adaptive versus Non-adaptive evolution
- Adaptive evolution has a predictable influence on average fitness in a population (it increases it), whereas non-adaptive evolution has a chance of increasing or decreasing average fitness
Principle of Mass Action (general case, not the one you might have covered in neurobiology)
- Although the idea of randomness implies unpredictability, in large populations over long periods of time, the aggregate effects of random events can be described mathematically, which makes it possible to predict the effect of drift on populations
- Thus, individual 14C atoms decay randomly but mass action makes it possible to predict when half of a population of them will have decayed
- One corollary to this is that population size is an important factor in making the predictions, making population size a factor that almost always has an impact
In an infinite population, the outcome of an event is exactly what its probability predicts:
- In an infinite population of fair coin tosses, 50% are heads and 50% are tails
- In a finite population, random chance means that the actual outcome can differ from the expected
- In a population of 100 coin tosses, 50 heads are expected but we would not feel that the universe had come unraveled if we got 53 heads (or 46 heads)
- In a population of 15 coin tosses, the predicted outcome (7.5 heads, 7.5 tails) is mathematically possible but practically impossible
- The difference between expectation and outcome is the error due to sampling (Sampling Error)
- Sampling is drawing a group of some given size out of a larger (theoretically, infinite) population
- If this does not seem true, then think of a sample of five coin tosses
- expected outcome (50% heads) would mean that you get 2.5 heads, an impossibility as you can get 2 heads or 3, but not 2.5
- Here the sample size is so small that the expected outcome from a known probability is not always even a possible outcome
- Sample size is an important factor in drift
- Small samples (populations) experience greater deviations from expectation
- Flip a coin 6 times and the expectation is 50% heads and the chance of 100% heads is 1.5% (=0.56)
- Flip a coin 20 times and the expectation is 50% heads but the chance of 100% heads is only 0.0001% (=0.520)
Because all populations are finite in size, in each generation, random chance will affect the proportion of each allele in the population
- Hardy-Weinberg predicts no change, just like we predict 50% heads, but random chance alters the actual outcome
- One measure of genetic variation applicable to Hardy-Weinberg situations is Heterozygosity (H)
- H for one locus is the fraction of individuals in a population that are heterozygotes at that locus
- for two alleles at one locus, then Hardy-Weinberg predicts that H = 2p(1-p) or = 2pq, when q = 1-p
- for three or more alleles, the number of heterozygotes goes up (if A is the number of alleles, then the number of possible heterozygotes is A(A-1)/2 - A (that works out to 3 for 3 alleles, 6 for 4 alleles, 10 for 5 alleles, etc.)
- in this case, it is quicker to figure out what Hardy-Weinberg predicts for the homozygotes (there are fewer of them to calculate)
- H = 1 - (sum of all homozygotes) or, for three alleles (for example), 1 - (p2+q2+r2 )
- H can also be the fraction of loci in an organism that are polymorphic - this is an unfortunate overlap in terms
- the book refers to this second meaning for heterozygosity in a sensible way - it calls it the proportion of polymorphic loci - but not all the literature is so exact.
- The overall effect of these "wandering" allele frequencies is that allele frequencies will wander until they are caught in one of two "traps"
- Fixation - when an allele's proportion hits 100%, no other alleles are left and we say that allele is "fixed" in the population
- Extinction - when an alleles proportion hits 0, it is lost from the population
- The direction of fluctuation (increase or decrease allele's frequency) is independent both of any changes that occurred previously (an increase is just as likely to be followed by a decrease as by an increase) and of the frequency of the allele (no matter the frequency, the chance of a random increase equal to the chance of a random decrease)
- This leads to a Random Walk of the allele through time, a walk that only ends with fixation or extinction of the allele
As you can see, without migration or mutation, the number of heterozygous loci in a population will decrease over time as one or other of the alleles at a polymorphic site (or locus) becomes fixed
Genetic Drift leads to a loss of genetic diversity
Some Impacts of Genetic Drift on Evolution
Linkage disequilibrium can arise through drift
- Two loci on the same chromosome with two alleles yields four combinations (or chromosome types): AB, Ab, aB and ab
- drift will cause fluctuations from the expected equal proportion of each combination (25% of each)
- Without recombination, drift will cause one of the four combinations to become fixed, which is extreme linkage disequilibrium
- With recombination, drift promotes disequilibrium and recombination breaks up allele combinations, so an equilibrium is expected
- The level of linkage disequilibrium is proportional to recombination rate (which depends on the physical map of the chromosome) and and the population size (which determines the rate of genetic drift)
Notice that population size is very important and deserves further consideration
- If you count the size of a population (census the population) and use that figure as N, you are assuming that all individuals have the same chance of contributing genes to the next generation, which is often not a fair assumption
- The actual number of breeding individuals is the Effective Population Size (Ne) - notice that Ne cannot be larger than N (the census size)
- Some factors that can reduce N (see Effective Population Size Worksheet for more on this subject)
- Variation in the number progeny - through chance or because natural selection is favoring some progeny over others
- Variation in the population sex ratio (away from 1:1) will reduce Ne
- Variation in population (census) size - when considering what happens over many generations, it is important to realize that chance events are more important during periods of low population numbers
- when population size varies through time, Ne is not the arithmetic mean of the population sized (this is the average we normally calculate by summing up the population sizes and dividing by the number of sizes summed) but is the harmonic mean (which is calculated as the reciprocal of the mean of the reciprocal population sizes)
- Note that the harmonic mean is always smaller than the arithmetic mean if there is any variation among population size usually closer to the minimum population size than the mean population size (it is always smaller than the mean)
- There are two extremes of the effect of variation in population size:
- Bottleneck Effect - when a population undergoes a crash, many alleles can be lost (think of a crash from 1,000 to 2 individuals - the large population may have had many, many alleles at a locus but the maximum number in the post-crash population is 4, as there were only two copies of the gene in each individual
- Founder Effect - essentially the same as the bottleneck but it is not a crash but the effect of a small group of individuals (sometimes just one pregnant female) founding a new population
Genetic Drift in a multi-population framework
Neutralist-Selectionist Controversy
- Selectionists saw adaptation and assumed that most variation at the genetic level had an effect on fitness (few neutral changes)
- Neutralists felt that most genetic variation had no effect on the phenotype or a very slight effect effectively neutral)
- Neutralists and selectionists agree that:
- beneficial mutations lead to adaptations, not random drift
- many mutations are harmful and are eliminated by natural selection
- Neutralists and selectionists disagree about
- The relative influence of natural selection versus genetic drift in evolution
- The proportion if effectively neutral mutations (less difference now than at the start of the controversy)
- The intensity of the controversy meant what it always means: both sides had data to back them up but not so much data that the other side had to give in
- the result of such controversies is always the same: both viewpoints are integrated into an overall understanding
- evolutionary biology is in the process of integrating neutralism with selectionism
- Most alleles are neutral (of equal fitness), some are harmful (lost from population due to natural selection), and some are beneficial (source of adaptations through natural selection)
- Neutrality is useful
- can be used to calibrate a "molecular clock" based on the rate at which new mutations arise and become fixed
- can be used to find ancient relatedness, as natural selection will promote convergence and parallel evolution, which produce homoplasies
Neutral mutation rate -u0
- u0 is the rate of mutations at a locus per gamete per generation
- most point mutations in a gene occur at unique sites (if they occur at the same site along the sequence, a new mutation would wipe out the presence of the previous mutation)
- some sites may be functionally constrained because changing the base pair would change the codon and the amino acid at that site in the protein
- if the amino acid is key to the function of the protein, any change may be harmful, not neutral (or, possibly helpful and still not neutral)
- e.g. - an amino acid that is important to the catalytic function of the protein
- The neutral mutation rate should equal the rate of fixation of mutations
- fixation rate will underestimate the mutation rate to the degree than new mutations occur at the same site
- estimation of can be done if you compare two sequences in two species and can use the fossil record to date the time of the split (or, if you have genes in two allopatric populations and you can date the time of the population split and assume no migration)
- the problem of multiple changes at one site again can invalidate this process and it gets worse as the time since the split increases
- it is also possible to correct for the likelihood of multiple hits
- Thus, at a locus, new mutations constantly occur and fixation (in the sense of an allele being "fixed" at a frequency of zero) constantly removes alleles, which will produce a balance over time
- At equilibrium, the proportion of the population that is heterozygous at a locus is a way to measure the balance of these forces and will depend on the neutral fixation rate and the population size (which determines the rate of loss of alleles due to fixation), These are related as:
Genetic Drift within a population will move the population toward fixation of one allele at any locus
Gene Flow of alleles from other population will bring in alleles and counter the effect of drift
- In populations where only drift and flow are occurring, these opposing forces produce an equilibrium where loss of alleles due to drift is balanced by gain of alleles through flow
F statistics can be used to predict the equilibrium point
- We know that FST is related to the variation of allele frequency among populations from the lecture on genetic variation
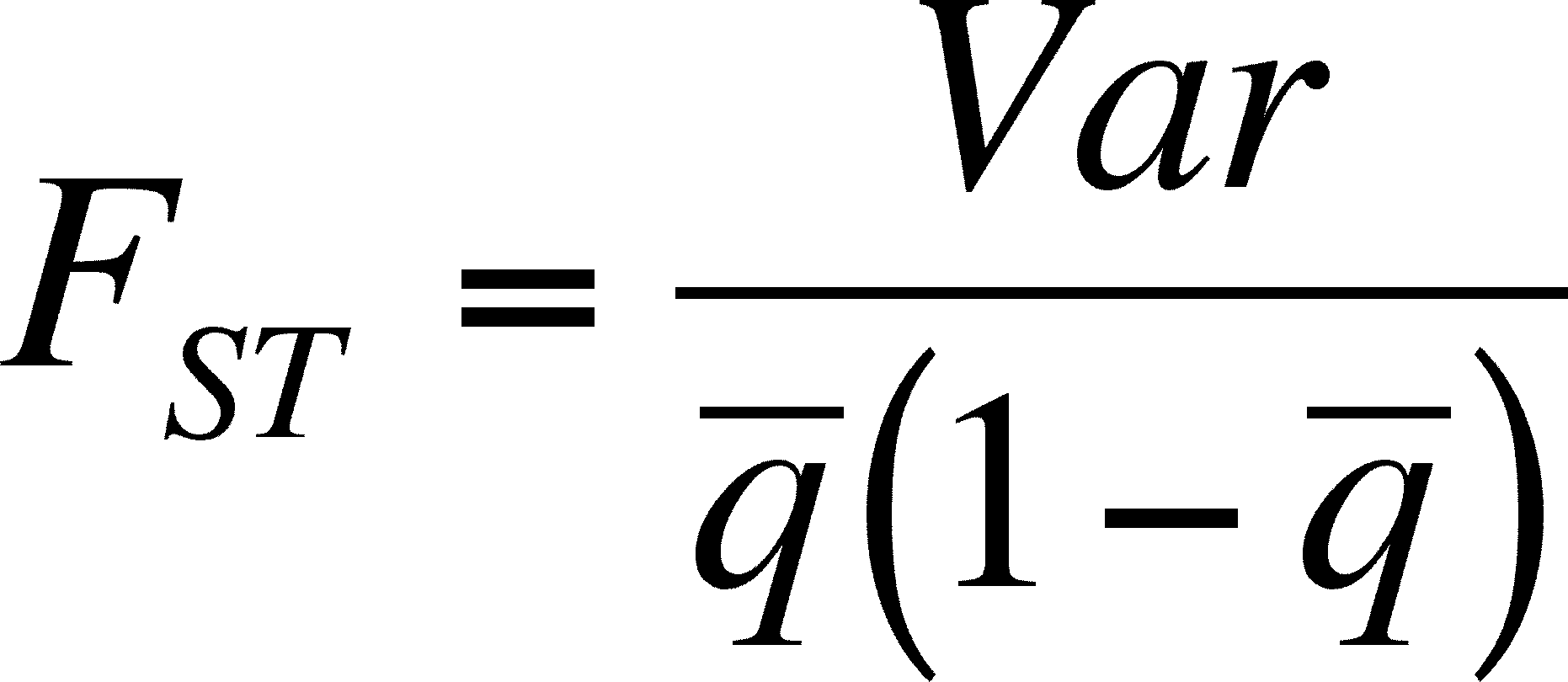
- FST is also approximately estimated with the formula below that relates it to population size and migration rate
- Migration rates are notoriously hard to estimate as they can vary greatly in time and it is often impossible to determine which are immigrants and which are not
- FST is often easier to measure as our ability to determine genotype becomes more accurate and facile
- So, FST is often a better way to determine gene flow than direct methods such as you learn in ecology class (mark-recapture, for instance), if (and it's a big if) you can assume that other factors (selection, assortative mating, mutation) are insignificant in comparison to drift and flow.
Coalescence, Drift and Population History
Coalescent Theory is based on the idea that genes present in today's populations have a single ancestor some time in the past
- This does not mean that the ancestral population was monomorphic, but that other lineages present then have since become extinct
- Coalescence models are simplest when the population is closed and there is no selection
- Drift produces extinction of the lineages at a predictable rate dependent on population size
- How many generations back it takes to coalesce to a single gene depends on population size
- Small populations coalesce more recently than large populations because drift will eliminate lineages sooner in small populations
- Genes coming from some unknown source will invalidate the methodology. Sources of such genes might be:
- intraspecific - from unknown or unsampled populations of the same species
- interspecific - new alleles arriving in a population by genetic exchange with another species - (Horizontal Gene Transfer)
- This theory allows for migration among populations but only among known populations with known allele frequencies
- Coalescent theory depends on the process of random loss of alleles due to genetic drift
We can use coalescence theory to predict the history of a population based on its current composition and size
Last updated March 21, 2011