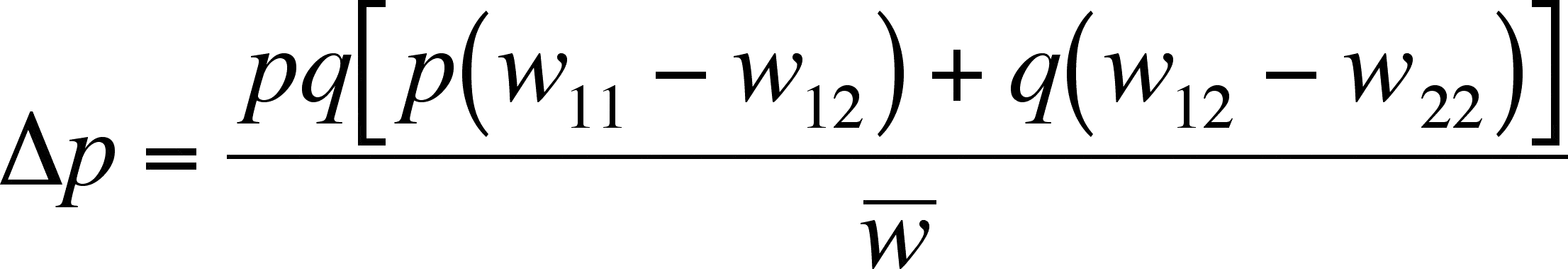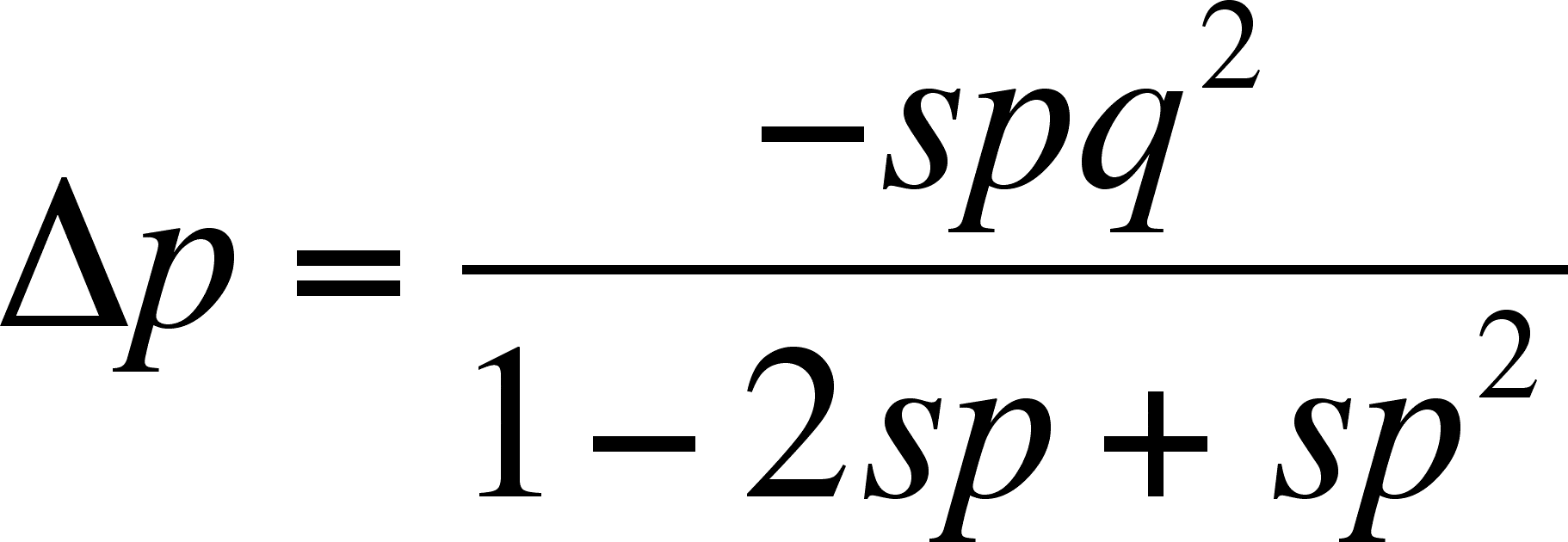|
BIOL 4160 Evolution Phil Ganter 301 Harned Hall 963-5782 |
Flower
of a Bunchberry, a Cornus species (the dogwoods) that
is not a tree and grows only inches high |
|
|
BIOL 4160 Evolution Phil Ganter 301 Harned Hall 963-5782 |
Flower
of a Bunchberry, a Cornus species (the dogwoods) that
is not a tree and grows only inches high |
|
Microevolution: Modelling the Natural Selection of Alleles
Link to a list of Specific Objectives for lectures
Back to:
Academic
Page |
Tennessee
State Home page |
BIOL4160 Page |
Ganter
home page |
Remember that natural selection is a mechanism of evolution, it is not evolution
Natural Selection can enhance, reduce, or maintain variability (in the last case, natural selection resists evolution!)
Fitness is the per capita rate of increase of some biological unit (we will see that this can be a locus, an individual, a deme, or even a species)
Absolute Fitness
- So, is there a measure that will combine all components
- Yes, there is. It is called r (or, as in the book, R (really R0), a related measure that is easier to explain), the intrinsic rate of natural increase
- When referring to a gene or lineage (can't apply to individuals, only the lineage to which they belong) the letter "m" is often substituted for "r", especially in the older literature
- r comes from ecology and we will not develop the theory to use it here, so we will conform to the book's use of R0, the symbol for the replacement rate
- For asexuals, is the number of individuals in the next generation for ever individual in the present generation
- if R0 is greater than 1, the population is growing
- if R0 equals 1, the population size is stationary
- if R0 is less than 1, the population is decreasing
- We can apply this idea to a portion of the population, say those with the A allele at locus A, so if A has an R = 1.4, then there are 1.4 copies of that allele in the next generation for each copy in the present generation
- R0 (or r) is a measure of absolute fitness because it literally indicates how many copies of a gene there will be in the next generation
- Absolute fitness is useful but can be hard to measure and hard to work with when modeling evolution
Relative Fitness
- Relative fitness is often preferred as a measure of fitness
- The fittest genotype (in absolute fitness terms) in a generation is the reference fitness
- All other genotypes will have a fitness less than the reference, and so they will be a fraction between 0 and 1 if we simply divide the absolute fitness of each genotype by the absolute fitness of the reference fitness, the maximum absolute fitness
- notice that the denominator is always at least as large as the numerator and will usually be larger, giving us the 0-to-1 range of relative fitnesses defined above
Mean relative fitness
- This is a measure of the difference between the actual population fitness and the theoretical maximum
- The rate of genetic change in a population due to selection will depend on the magnitude of the difference between actual and maximal fitness
- calculated by multiplying the relative fitness of each genotype by its frequency in the population, then summing up all of the products
- If absolute fitness (R0) of A1A1 is 1.6, is 1.36 for A1A2 and 1.2 for A2A2,
- then the relative fitness (Wi) of A1A1 is 1.0, is 0.85 for A1A2 and 0.75 for A2A2
- Now suppose that half of the population is A1A1 and the other half is equally split between A1A2 and A2A2
- Then the mean fitness of the population (W-bar) is
- (0,5 * 1.0) + (0,85 * 0.85) + (0.25 * 0.75) = 0.5 + 0.2125 + 0.1875 = 0.9
Coefficient of Selection (often s)
- measures the selective disadvantage of a genotype relative to the most fit genotype
- calculated as s = 1.0 - W (using the relative fitness of a particular genotype)
One additional and important advantage of using relative fitnesses
- The rate of genetic change under selection depends not on the absolute fitnesses but on the relative fitnesses
- So, it doesn't matter if the R0's are 1.6, 1.36, and 1.2 or 8, 6.8 and 6 or 0.5, 0.425, and 0.375 (this last for a declining population), evolution will proceed in the same manner and speed because the relative fitness of all three scenarios is 1.0, 0.85, and 0.75.
- Note that it does not matter if the population is growing or not, evolution will proceed as expected
Genotypes Frequency of Genotypes at Birth Fitness of Genotypes
A1 dominant and advantageous (A2 disadvantageous)
- First, and for all of the equations below, we will express the relative fitnesses in terms of selection coefficients and then present the algebraic simplification that results from substituting them into the general equation above (all five cases are simplifications to predict the outcome in specific situations)
Fitness
- This says that p will increase (the right side of the equation is positive and so delta-p is positive), which makes sense as A1 is the most fit of the two alleles
A1 dominant but A2 selectively advantageous
Fitness
- Here delta-p is negative (look at the right side) and so A1 is being lost from the population, as is should if A2 is more fit than A1
Incomplete Dominance with the heterozygote fitness between the advantageous dominant homozygote and the disadvantageous recessive heterozygote
- In this case, the heterozygote must have a higher fitness than the homozygous recessive genotype and one way to do that is to multiply s by a second fraction, h
- h will increase w because, as h is between 0 and 1, the product of h and s will be smaller than s and this smaller product is subtracted from 1
Fitness
- This equation predicts that the change in p is always positive, so the endpoint arrives when A1 is fixed in the population
- Note that this equation differs from the text's equation (equation A3 on page 274). I could not derive the equation presented in the text but, when I graphed the equation, it did not behave as described there. So, I am presenting my derivation, which does behave as predicted (and as makes sense because w11 is the fittest genotype and should move A1 to fixation)
Incomplete Dominance with Heterozygote advantage
- The heterozygote has the fittest genotype but we will allow each of the homozygotes their own selection coefficient
Fitness
- The change in p will depend on its frequency: it will be positive below a particular value of p and negative when p is over that value (change = 0 when p is at that value)
- This scenario produces a Balanced Polymorphism, a stable equilibrium (stable because when not at the equilibrium point the value of p moves toward equilibrium so that the system returns to equilibrium)
- This situation is called Heterosis
Incomplete Dominance with Heterozygote disadvantage
Fitness
- The outcome here is also an equilibrium but an unstable equilibrium
- When the value of p is not at the equilibrium point the change in p will not move it toward the equilibrium point but away from it (toward either fixation or loss of the A1 allele, depending on the value of p)
A special case - A1 is dominant and A2 is a recessive that is lethal when homozygous (harmless when heterozygous due to the dominance effect)
- In this case, we see that dominance can protect the heterozygote but every generation, the homozygous recessive individuals are lost before they can reproduce (or even before they can develop)
Fitness
- Notice something about the equation above
- As q, the frequency of the lethal allele (A2) decreases, as it should (after all, its lethal in the homozygous condition), the rate of change for p is smaller and smaller (it depends on q2, which is a smaller and smaller numerator)
- Thus, the rate of loss of the lethal slows to a trickle and it persists in the population for many generations
- This makes sense because, when there are very few A alleles present, the chance of an individual carrying A mating with another rare carrier of A is very small and that is the only way to get homozygous individuals that will be lost
![]()
Detecting Selection in the Gene
Background:
Consequences:
Last updated March 24, 2010