|
BIOL 5130
Evolution Phil Ganter 301 Harned Hall 963-5782 |
|
Eyespot on the wing of a Polyphemus moth |
|
|
BIOL 5130
Evolution Phil Ganter 301 Harned Hall 963-5782 |
|
Eyespot on the wing of a Polyphemus moth |
|
Brief History of Life
Lecture 01
Back to:
Paleontology is the study of life through the examination of fossils
Palynology is closely related (perhaps even a subdiscipline) that studies contemporary and fossil palynomorphs (organic microfossils)
- primarily pollen but also including:
- spores, dinoflagellate cysts but excluding acid-soluble fossils (carbonate and siliceous fossils)
- chitinozoans - odd flask-shaped organic fossils from the Ordovician and Devonian seas (tests? egg cases? who knows!) with lots of ornamentation on their surfaces
- scolecodonts - the hard parts of polychaete jaws, common in many marine sediments
- acritarchs are small organic fossils that aren't classifiable as part of any other recognized type of palynomorph
- Palynologists go beyond organisms to the study of organic molecule remains, including particulate organic matter (POM) that settles out of the water column and becomes part of sedimentary rock
- kerogen is a mixture of organic compounds found in sediments and sedimentary rock (can be large molecules, on the order of
- these studies have resulted in the processing of sedimentary rocks (oil shales) to obtain gas and petroleum
Taphonomy is the study of organic decay (what happens after death) and includes the study of fossil formation
- Necrology is the study of the death of organisms (are they prey? is the fossil merely a part of an organism that regularly dies and is cast off, like deciduous leaves?)
- Biostratinomy is study of the process of including organic remains into the sediment, the burial process
- Diagenesis is the process of lithification of organic remains (to lithify is to turn into rock) and includes all changes that take place after an organism is buried in the sediment
How do we gather data on the history of life?
Fossils
Any trace of past life can be considered a Fossil (something dug up)
- Remains of hard parts of organism (shells of anything from microbes to large bones and mollusc shells)
- Petrification - the replacement of organic structures by mineral crystals that reflect the organization of the replaced structures
- Trace fossils
- Tracks in sediments
- Compressed impressions left by soft body parts in sedimentary rock
- Burrows left in sediments by worms and other invertebrates
- Coprolites or Coproliths - fossilized feces
- Middens - collections of food materials, nesting materials, etc. formed by some rodents (pack rats) - these are not very old but do provide a detailed recent history
Macrofossils - as the name implies, these are large fossils detected by the naked eye
Microfossils - the remains of microbes or parts or structures of larger organisms that require the use of a microscope for study
- shells (CaCO3 or SiO2) of protists, pollen of plants, cysts of Sarcodines and other protists, Conodont (early vertebrate) "teeth" are all included
- many bacteria have left traces in rocks and these constitute the oldest fossils, over 3.5 billion years old
- Stromatolites (mushroom-shaped calcareous deposits from cyanobacterial activity) are the oldest
- old microfossils are controversial as it is hard to rule out formation of the fossil by abiotic processes
Dating Rocks
See next section on Geological Time for relative ages
Absolute ageing is done through Radiometric Dating
- Radioisotopes (Radionuclides) decay into Daughter nuclides, often radioactive themselves, and may form a several step "lineage" of new atoms from the original
- The chance that this decay will happen is set by nuclear events and is not affected by pressure, temperature, etc. experienced on the Earth
- Thus, the time for half of the radionuclide atoms to spontaneously decay is set by the nature of the isotope and is a constant for a particular radionuclide (this is the Half Life of the radionuclide)
- Half-lives range from milliseconds (some man-made elements) to billions of years (704 million for 235U/207Pb, 1.25 billion for 40K/40Ar, 4.5 billion for 238U/206Pb, 48.8 billion for 87Rb/87Sr and 106 billion for 147Samarium/143Neodymium) for different radionuclides
- Thus, if you know the current concentration of the radioisotope and its daughter nuclide, you can work out how many half-lives ago the rock contained all radionuclide and no daughter atoms
- Note that you have to assume no daughter nuclides were present at the start or you have to be able to correct for a known starting concentration
- The equation for this calculation is:
Time = 1/
ln(1+P/D)
- Where
- Some things to know here:
- This type of dating assumes that the countdown begins when the crystal is formed - when igneous rock forms from liquid magma and sets the amount of radionuclide in the material
- No more radionuclide can enter the material or the date will be wrong
- The shorter the cooling time, the more accurate the dating
- Sedimentary rock can't be dated with this technique as the crystals that form it were formed when the igneous rock that weathered to form the sediments being compressed into rock was formed
- Dating would tell you when the parent rock was formed, not the sedimentary rock
- There are some crystals that form as sedimentary rock is formed on the ocean bottom, and these can be directly dated
- Since much of the rock containing fossils is sedimentary, fossils can't be dated directly this way
Radiocarbon dating can be used to date recent fossils that contain some of the original animal or plant
- Radiocarbon dating has a problem: the daughter nuclide is 12C, the common isotope of carbon, and the fossil is probably full of 12C atoms, so which are daughter atoms and which were not produced in the fossil by radioactive decay?
- Radiocarbon dating is slightly different for this reason
- Cosmic rays and natural radioactive decay both form radioactive isotopes from non-radioactive isotopes
- e.g. cosmic rays strike 14N and transform it into 14C, which decays into 12C
- When alive, organisms exchange carbon atoms continually through respiration and assimilation, so organisms reflect the environmental level of 14C
- After death, no more exchange so 14C levels begin to decline as the 14C decays into 12C
- For radiocarbon dating , you know the starting and final concentration of 14C and use that to determine the number of half-lives since the death of the organism
- Another problem for radiocarbon dating:
- the half-life of 14C is only 5730 years and radiocarbon dating is only good for the last 70,000 years (about 12 half lives, which will reduce the 14C concentration to 1/4192 of the original)
- Thus, radiocarbon dating is more useful for anthropologists and some palynologists than for paleontologists
Geomagnetic Polarity Reversals
- Before igneous rock solidifies or as sediments are deposited, ferromagnetic particles are oriented by their interaction with the global magnetic field
- When solidified, these particles give the rocks containing them a magnetic polarity in line with the Earth's magnetic polarity
- Discovery of polarity reversals revealed by striping in rock layers (stripes are formed by zones of reversed polarity) led to the discovery that the Earth's magnetic poles reverse at irregular intervals
- The reversals are proceeded by a reduction in overall magnetic field strength
- The reasons for reversals are not known but it is known that the magnetic field of the Sun also reverses at much shorter intervals (every sunspot cycle is accompanied by a magnetic reversal) although, because the reversals in the sun's polarity are associated with periods of magnetic field strength and the Earth's reversals with magnetic field weakness, it is not known if they are caused by the same phenomenon
- Some speculate that the reversals are the outcome of inhomogeneities in the Earths molten core, which rotates and acts as a dynamo in generating the field and its polarity
- Some speculate that external influences on the Earth's core are the reason (tectonic events or even extraterrestrial impacts)
- The discovery of magnetic striping, once appreciated (it was discovered by a Japanese researcher, Motonari Matuyama, in the 1920's but did not have its impact until the early 1960's), it revolutionized our understanding of geology and lead to the current idea of continental drift and tectonic plates
- By carefully measuring and documenting the striping, we now have a Geomagnetic Time Scale with which to calibrate rock ages from different parts of the world
- Some time periods have many reversals, which can occur within tens of thousands of years
- These can be very useful for determining if two fossils are contemporaneous
- Some periods have long intervals between reversals of tens of millions of years (these are called Superchrons, such as the Cretaceous Superchron)
Molecular Clocks
- Radiometric dating is our first method of absolute dating and is considered the gold standard (even the Geomagnetic Time Scale is based on radiometric dating)
- In 1962, E. Zukerkandl and Linus B. Pauling (he of the alpha helix, of vitamin C as a means of avoiding the flu, and of nuclear disarmament) "Molecular disease, evolution, and genetic heterogeneity", in Kasha, M. and Pullman, B (editors): Horizons in Biochemistry. Academic Press, New York, 189–225, noticed that the number of changes in the sequence of various haemoglobins corresponded to the proposed divergence times of the taxa in which they were found
- They proposed that the change was a regular occurrence and that, due to its regularity, could be a "molecular clock" in which each change was a tick indicating the passage of a constant amount of time
- In 1967, V. M. Sarich and A. C. Wilson ("Immunological time scale for hominid evolution". Science 158 (3805): 1200-1203) and 1968, Motoo Kimura ("Evolutionary rate at the molecular level". Nature 217: 624-626) proposed that neutral DNA changes constituted a molecular clock
- Neutral sequence substitution happen as the outcome of a spontaneous mutation that is fixed by stochastic events and, given the rarity of mutation and reasonably constant population sizes, should happen at a reasonably constant rate
- Thus, a molecular distance is converted to a divergence time through the assumption of rate constancy
- Mutations fixed by selection are not governed by the constant force of random chance and may deviate from a clock-like regularity
- For instance, in some regions of DNA with a constant function, stabilizing selection might resist sequence change and slow the clock, whereas directional selection might speed the rate of substitution in regions undergoing a shift in function
- Molecular clocks need to be calibrated
- Must have a fixed time in the past, which is usually a well-dated fossil ancestor, assumed to have the hypothetical ancestor's gene sequence
- Attempts to do so have met with both successes (H. Ochman and A. C. Wilson. 1987. "Evolution in bacteria: evidence for a universal substitution rate in cellular genomes.". Journal of Molecular Evolution: 74-86.) and failures (E. J. P. Douzery, F. Delsuc, M. J. Stanhope, and D. Huchon. 2003. "Local molecular clocks in three nuclear genes: divergence times for rodents and other mammals, and incompatibility among fossil calibrations". Journal of Molecular Evolution 57: S201-S213)
- Recent work has attempted to allow heterogeneity in evolutionary rate, a kind of (as one article puts it) "relaxed molecular clock" so that a strict assumption of rate constancy is not needed but one can obtain an estimate of divergence time (many methods also attempt to calculate confidence limits about the estimate as well)
- As expected, molecular clock estimates often agree with fossil evidence but there are significant disagreements between fossil and molecular clock estimates
- The most interesting is the controversy over the meaning of the "Cambrian Explosion"
- Since this marks the debut of many phyla in the fossil record (most of which do not last long), it has served as a nexus of research for those interested in the history of life
- If it is an explosion, then multicellular animals originated in the Proterozoic, perhaps as long as 2.5 bya and limped along as a small, homogenous group until the Cambrian, about 0,55 bya, when they suddenly increased in diversity beyond the diversity we know today and from that time, we have lost phyla while some phyla have sub-diversified
- this is the dominant hypothesis and one held dear by paleontologists, many of whom ascribe to a view of macroevolution called "Punctuated Equilibrium"
- Punctuated Equilibrium (in a sentence) is the idea that diversification (at the level detectable through fossils) occurs not gradually (as Darwin postulated) but that there are long periods of "stasis" during which diversity will remain unchanged or even fall as forms are lost and that stasis is "punctuated" by sudden bursts of diversification
- It is not exactly known why some periods are static and some are punctuated but most explanations come down to either evolutionary novelties opening new habitat for exploitation (like the novel feeding methods found among the cichlid fish of Africa's rift lakes) or through the loss of some lineages opening habitat into which remaining groups to diversify (like the explosion of the mammals after the loss of the large dinosaurs).
- Molecular biologists, looking at divergence for genes found in all (or almost all) living multicellular organisms, date the divergence deeper than the Proterozoic-Cambrian boundary, indicating that the Cambrian explosion is an explosion in fossils (an undeniable fact) but that the animals had originated before the explosion
- As molecular dating pushes the origin of today's animal phyla deeper into the past, and breaking the divergences into more than a single explosion, the champions of Darwinian gradualism take heart
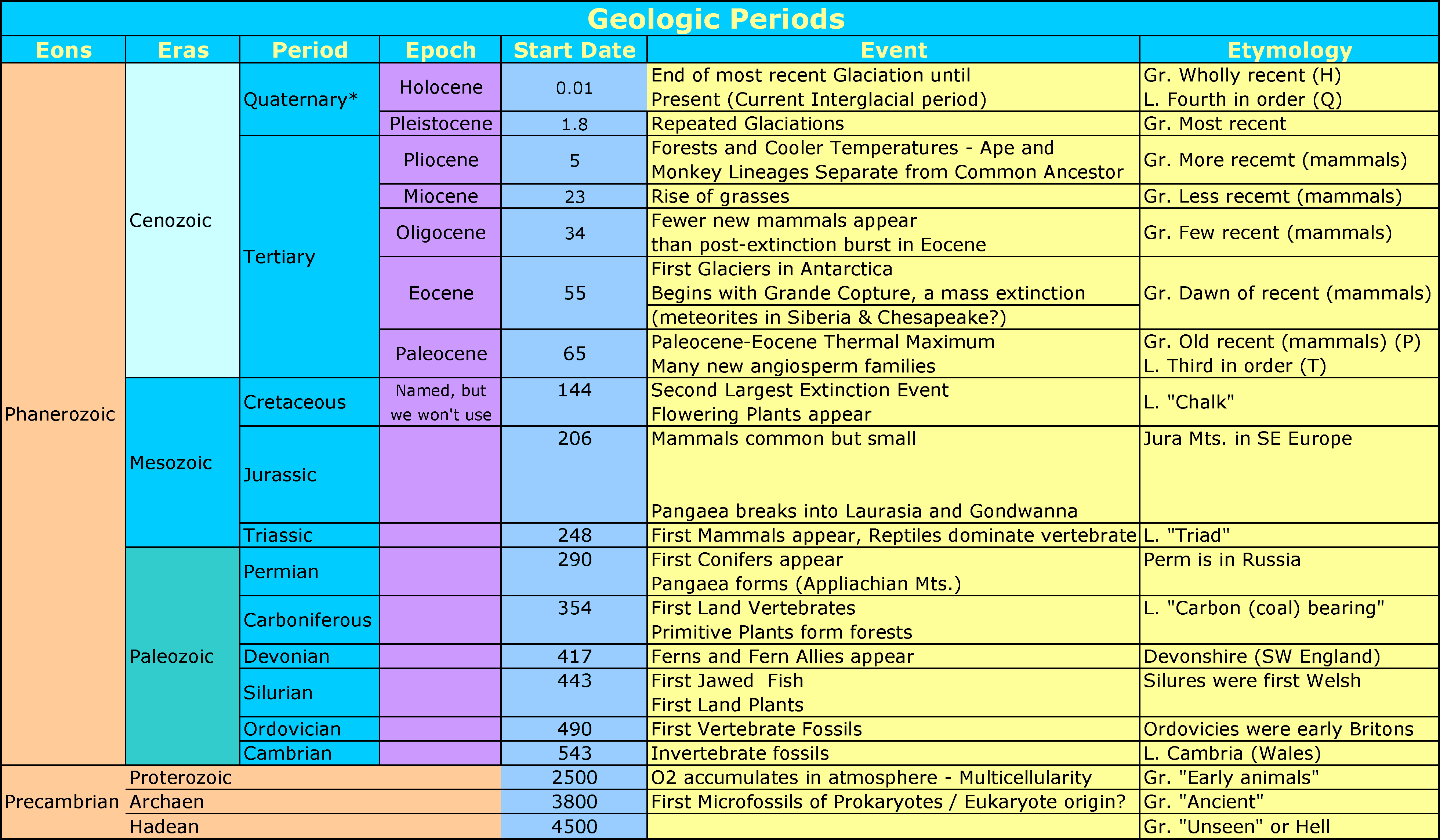
Earth's History and Geological Time
*A recent proposal has the Quaternary as an era after the Cenozoic and the Cenozoic divided into the Paleogene and Neogene periods (some of the current Quaternary is part of the Neogene) but it is not yet officially accepted (as of the date below)
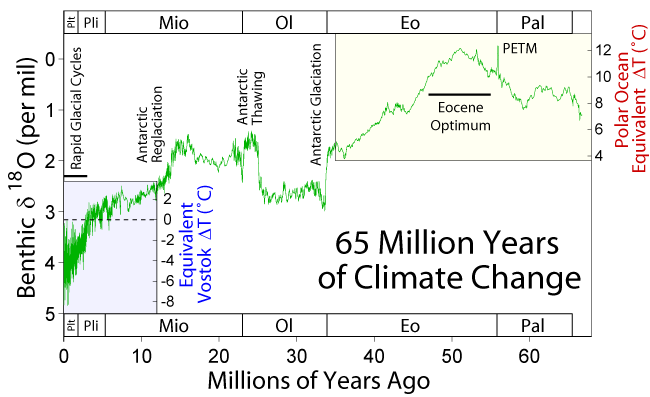
Graphic from Wikipedia (see page on Paleocene-Eocene Thermal Maximum)
Plate Tectonics and Earth's History
Surface is divided into a series of plates that move independently and can collide, split apart, and overlap
Origin of Life
Early Evolution (before the Age of Macrofossils)
Cambrian Explosion and the Age of Macrofossils
Invasion of the Land
Chordate Evolution
Chordate Bauplan
Vertebrates
Hominids
Macroevolution and Microevolution
nPr = nr
Blair
Last updated April 7, 2008