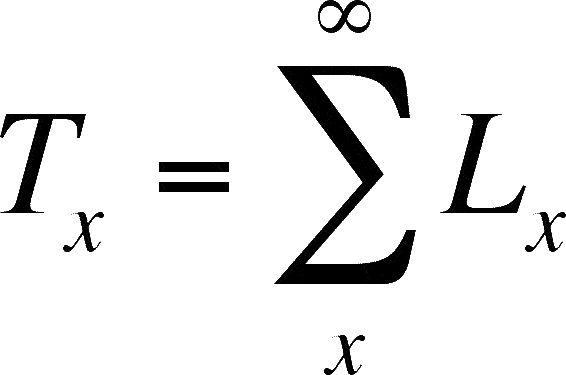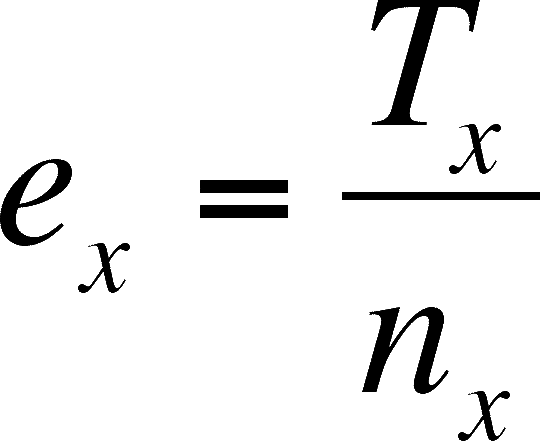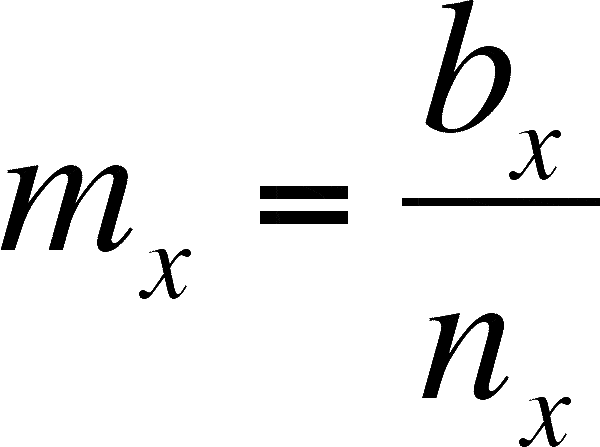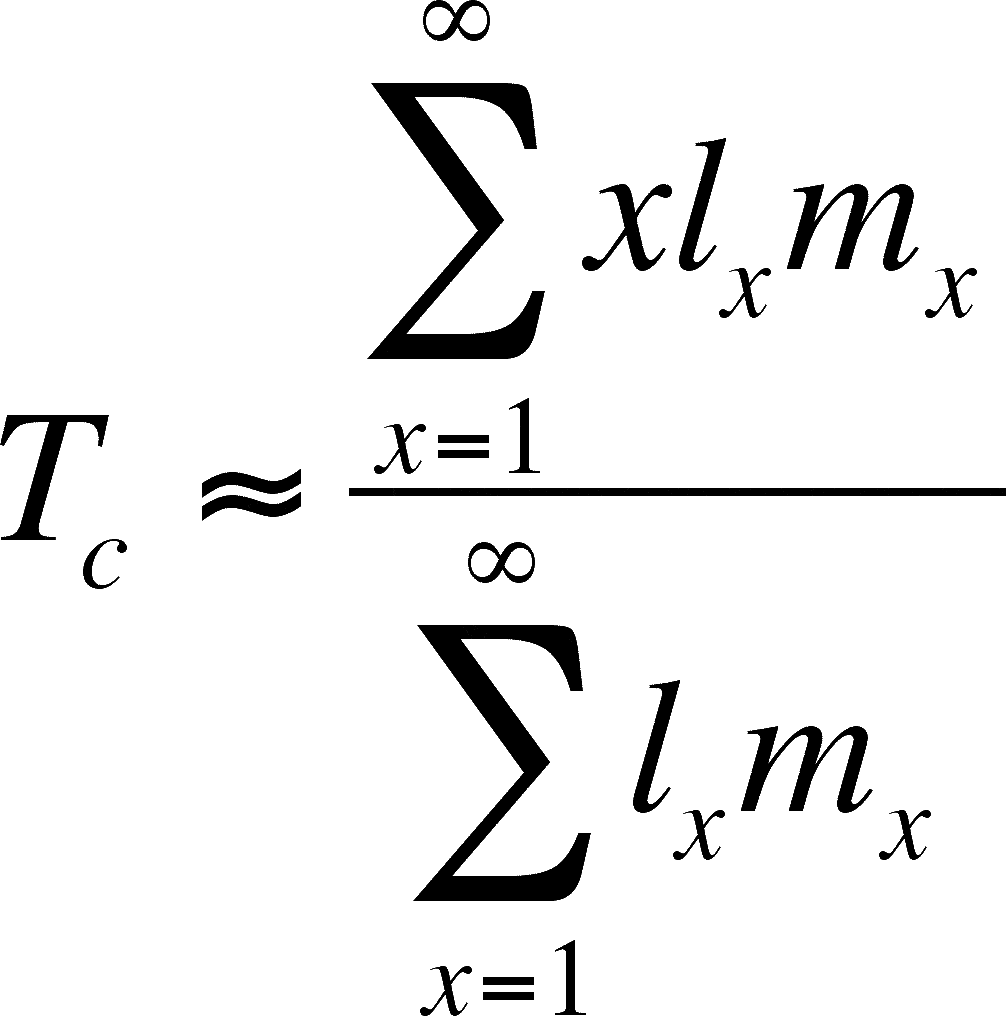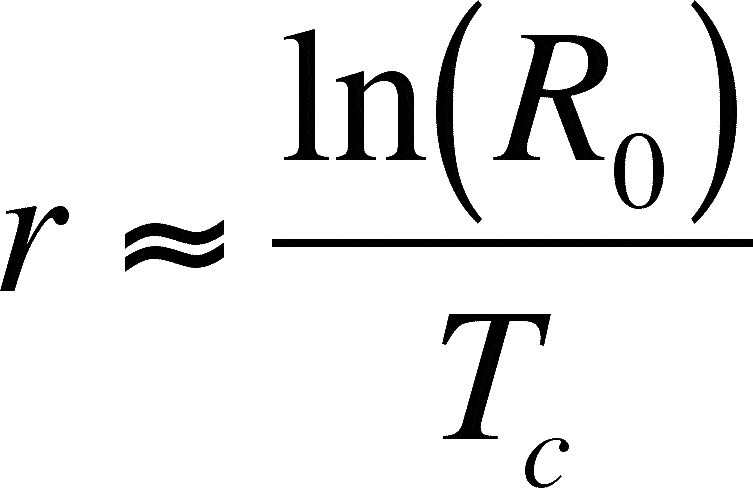|
BIOL 4120 Principles of Ecology Phil Ganter 302 Harned Hall 963-5782 |
| |
BIOL 4120 Principles of Ecology Phil Ganter 302 Harned Hall 963-5782 |
Chapter 6 Population Growth
Back to:
Sections:
Descriptions of Populations:
Demography is the science of life tables
Life Tables are accounting of births and death in a population
Usually the life span of an individual is broken into stages (egg, larvae, etc.) or by age intervals or classes (0-5 years, 5-10, etc.)
Cohort - all of the individuals born at the same time
x = the interval
nx = the number alive at the START OF THE INTERVAL
lx = age specific survivorship -- fraction of cohort alive at the START OF THE INTERVAL
dx = age specific mortality -- number dying in the interval
qx = age specific mortality rate -- death rate for individuals of a specific age
ex = age specific life expectancy - number of years left for an organism of a particular age. IT IS NOT ALWAYS MAXIMAL AT BIRTH!!
bx = age specific natality -- number born in the interval
mx = age specific birth rate - the per female rate of offspring production for females of a particular age.
R0 = net replacement rate = the number of females alive in the next generation for each female alive in the present generation.
T = generation time -- the average time between a birth of a member of the cohort and the birth of a member's offspring
Age specific means that the parameter varies with the individual's age
By convention, the first age interval has the notation 0, so the number in the cohort at the start is n0.
Another convention (especially for human life tables) is to multiply the number in the lx column by 1000 (if it is actually 0.562, then it reads 562).
However, if you wish to calculate the net replacement rate (equation given below), then you must either convert lx back to a proportion or you will be calculating the number of females in the next generation per 1000 females alive in the present generation!
If we plot log (nx) versus age, we can see where most of the mortality occurs by where the numbers drop
To calculate ex, we will need to tack on another pair of columns to make it easier to do. This column is Lx, the average number alive in interval x. This is simple to do:
From this column, we can get Tx, which is the sum of all of the Lx values from x until the age that the last member of the cohort dies (this is symbolized by L°).
Life expectancy at age x is simply the average number of years lived by members of the cohort that are age x.
Net replacement rate is a measure of relative growth. It is the number of females alive in generation x+1 for each female alive in generation x. If it is greater than 1, the population is growing as there are always more females alive in the next generation than in the present. If less than 1, the population is decreasing (less than 1 female in the next generation per each alive in the present generation), and a net replacement rate of 1 means each female is exactly replaced in the next generation, so the population is stable, neither growing nor declining.
If we eliminate the males and add a column containing births, then we can calculate the net replacement rate. First, we must calculate the per capita birth rate (mx) for females of age x (females only!)
The net replacement rate is the sum of age-specific birth rates times the age specific survivorship. This means that an age class's contribution to the replacement rate must take account of the probability of surviving to that age.
The net replacement rate is a measure of population growth rate.
If it is 1, then the population in neither growing or declining in size (a stable population).
If it is less than 1, the population is declining by that proportion each generation (if it is 0.5, then the population will be half as big each generation).
A value over 1 indicates a growing population (a value of 2 is a population that doubles each generation).
Assume that the net replacement rate is near 1. If so, we approximate the generation time as:
The approximation is due to the discrete nature of a life table, versus the continuous nature of time (an integral approach would get the exact figure)
Deterministic Models of Population Growth:
A model is a description of a natural phenomenon
Can be
Can be:
Geometric (discrete generations) and Exponential (overlapping generations) Population Growth
Populations with overlapping generations (us!) use continuous equations
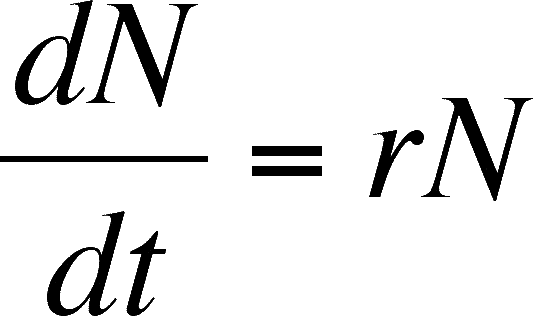
The instantaneous rate of increase of a population (dN/dt) is the result
For more on this topic, go to Modeling Exponential Growth page
In the last equation above, the rate of population increase simply increases with N
Logistic growth is based on the idea of a Carrying Capacity (K) for any environment, a population size:
- Above which the population decreases
- Below which the population increases
- When the population size is at the carrying capacity, then no growth occurs
- the change in dN/dt with a change in N is linear
- a graph with N as the X-axis and dN/dt on the Y-axis would be a straight line
- for animals with complex life histories and mating systems, this assumption is not always met
- social systems which entail some individuals not breeding when they could obviously violate this assumption
- there are no lags in the timing of the change in dN/dt with any change in N
- lags are common when life histories are complex, as in holometabolous insects
- larvae live in different environment and a change in larval density may not have an effect on egg-laying until after they pupate and become adults
- constant K (constant environment over space and time)
- K is likely to change over both space and time
- constant r (all individuals equally fit)
- often only a portion of a population breed, the rest may be helpers or may not have access to enough resources to breed
- no migration
- dispersal can be important, even keeping a population from going extinct when the net replacement rate is below 1
For more on the logistic, including a derivation and some problems, go to Modeling Density-Dependent Growth
Usually based on discrete models, but make allowance for the fact the things vary in the real world
Predictions of population size are couched as probability distributions
In general, the longer in the future you try to predict population size, the larger the range of possible sizes
Size and generation time are positively correlated (as one increases, so does the other)
Longer generation times are associated with lower intrinsic rates of growth
Demography, Life Tables, Cohort, Age specific, age specific survivorship, age specific mortality, age specific mortality rate, age specific life expectancy, age specific birth rate, Type I, Type II, and Type III Mortality curves, Net Replacement Rate (R0), Generation Time (Tc), model, verbal, pictorial and mathematical models, Deterministic and Stochastic models of population growth, Exponential (= Geometric) Growth, overlapping generations, continuous equations, intrinsic rate of natural increase (r), instantaneous rate of increase of a population (dN/dt), Carrying Capacity (K), sigmoid curve, stable equilibrium, logistic growth model, Time lags, chaos, deterministic and stochastic models
Last updated September 9, 2006